Thank you, it's cool. Used for the first time because only then was the right drug. Very revealing detail: ordered Express shipping to get the order within 3 hours because didn't know how to get the order faster https://africarx.co.za/buy-levitra-south-africa.html The drug is authentic exactly. Consistent with the stated prices the Staff is knowledgeable.
Variability of cuban and international populations of <emphasis type="italic">alternaria solani</emphasis> from different hosts and localities: aflp genetic analysis
European Journal of Plant Pathology 110: 399–409, 2004. 2004 Kluwer Academic Publishers. Printed in the Netherlands. Variability of Cuban and international populations of Alternaria solani from different hosts and localities: AFLP genetic analysis
Sim´on P´erez Mart´ınez1, Rod Snowdon2 and J¨orn Pons-K¨uhnemann2,∗1Dpto, Fitopatolog´ıa-Centro Nacional de Sanidad Agropecuaria (CENSA), Carretera de Tapaste yAutopista Nacional, San J. de las Lajas, Habana, Cuba (E-mail: sperez@censa.edu.cu);2Institut f¨ur Pflanzenbau und Pflanzenz¨uchtung, Justus-Liebig-Universit¨at, Heinrich-Buff-Ring 26-32,D-35392 Giessen, Germany; ∗Author for correspondence (Phone: +49 641 37542; Fax: +49 641 37549;E-mail: joern.pons@agrar.uni-giessen.de)Key words: Alternaria solani, genetic variability, AFLP analysis, host specificity
Abstract
As causal agent of early blight disease in tomato and potato, Alternaria solani is an internationally importanthorticultural pathogen. Genetic variability was surveyed by amplified fragment length polymorphism analysis ina total of 112 isolates from potato and tomato, representing pathogen populations from different Cuban provincestogether with isolates from the USA, Brazil, Turkey, Greece and China. Also included in the analysis were isolatesfrom catenulated Alternaria spp. from Brazil, Canada, Greece and Russia, along with single isolates of Alternariaporri, Alternaria alternata and a Curvularia sp. UPGMA clustering revealed a differentiation between the isolatesof A. solani and all other species with the exception of A. porri which could not be distinguished from A. solani. Among the isolates of A. solani, two distinct subclusters were formed, with high genetic significance revealed bybootstrapping, corresponding to a general subdivision based on the respective solanaceous host. The results arediscussed with regard to potential host specificity of A. solani on tomato and potato, and in terms of the comparativecontributions of regional and international genetic variability in populations of this ubiquitous plant pathogen. Introduction
Several studies have shown that A. solani isolates
differ in morphology, physiology, pathogenicity,
Early blight disease caused by Alternaria solani is one
genetic makeup and cultural properties. Indeed, iso-
of the most important fungal diseases of tomato and
lates can vary so much in their cultural characteristics
potato. This pathogen is also the causal agent of col-
that it is possible to find almost as many morpho-
lar rot in tomato. A. solani is a well-known pathogen
types as the number of isolates tested (Rotem, 1966).
of the genus Alternaria and is considered a good
Isolates also vary in the production of phytotoxins.
example for the world-wide distribution of a species
However, no correlation was found between the vir-
(Rotem, 1994). The fungus was described for the first
ulence of specific strains and their ability to produce
time in potato late in the 19th century (Ellis and Martin,
toxins (Stancheva, 1989). In some cases, strains
1882) and attacks on potato and tomato during the
from leaf and potato tubers differed in their ability
first three decades of the last century, on practically all
to attack potato leaves and tubers, but no correla-
the continents, were described by Neergaard (1945).
tion was observed between potato isolates and their
The first report of A. solani in Cuba was in 1918,
pathogenicity on leaves and tubers (Bonde, 1929;
at which point early blight in tomato was among
the most important crop diseases in Havana (Bruner,
Genetic analysis with isozymes (Petrunak and
Christ, 1992), RAPD (Sharma and Tewari, 1998;
Weir et al., 1998; Roberts et al., 2000) and RFLP
Materials and methods
markers (Adachi et al., 1996; Aradhya et al., 2001) hasshown great variability between and within Alternaria
species. A significant genetic distance was observedbetween isolates of A. solani from tomato and potato
Isolates of A. solani were collected from typical symp-
(Weir et al., 1998), suggesting the possibility of
toms of tomato and potato early blight plants in dif-
a pathogenic specialization on solanaceous hosts.
ferent provinces in Cuba (Table 1). Additionally, inter-
Generally, at the species level, the genetic variability
national isolates were provided by other researchers:
corresponds to variation in morphological characteri-
Ten isolates originated from Turkey, eight from Brazil,
stics (Roberts et al., 2000; Sharma and Tewari, 1998) in
four from Greece, eight from the USA and one from
Alternaria populations of fruits and crucifers, respec-
China. One isolate of Alternaria alternata, one of
tively. On the other hand, correlation of genetic markers
Alternaria porri, seven of catenulated Alternaria spp.
with host, geographic origin or resistance to fungi-
and one of Curvularia sp. were included in the analysis.
cides was not always evident (Adachi et al., 1996;
Origins and hosts of the isolates are listed in Table 1.
Morris et al., 2000; Aradhya et al., 2001). A high levelof genetic differentiation by host and/or pathogeni-
Table 1. Hosts and origins of fungal isolates used in AFLP
city, consistent with a hypothesis of specialization,
has been observed in Alternaria spp. from Citrus(Peever et al., 1999). In Cuba, the study of the vari-
ability of the causal agent of early blight has been
limited to tomato, for which 12 races were found in
Havana province (Izquierdo, 1981). More recently, we
observed pathogenic (Mart´ınez et al., 2002) and genetic
diversity in some Cuban isolates using RAPD analysis
Amplified fragment length polymorphisms (AFLP;
Vos et al., 1995) represent a powerful, highly repro-
ducible, PCR-based DNA-fingerprinting technique for
DNA of any origin or complexity. Because a large
number of polymorphic loci can be investigated in a
single experiment, the AFLP technique has become
one of the major methods of choice for studies of
genetic diversity, particularly in species where mark-
ers requiring genomic sequence information are not
available. The robustness of the AFLP procedure on
fungal genomic DNA was corroborated by Pei and
Ruiz (2000). The highly polymorphic nature of AFLP
markers make them especially useful for differentiat-
ing clonal lineages of fungi that reproduce asexually
(McDonald, 1997). AFLP markers have been used to
study genetic diversity in 18 isolates of Alternariabrassicicola (Bock et al., 2002). However, a large-
scale international survey of genetic variability in
Alternaria has not been published. The objectives of
the present study were to use AFLP analysis to examine
genetic differentiation within and between populations
from diverse tomato and potato growing regions in
Cuba and other countries, and to determine whether
the influence of the host could be observed at the
Curvularia Curvularia∗∗ Tomato 1999
Letter codes of the Cuban isolates are explained in the key to
Figure 2. Total numbers of isolates are given in brackets. A
double asterisk indicates monosporic isolates, whereas single aster-
isks show isolates that were not monosporic but where spores
were observed during the experiments. No spore production wasobserved in isolates without asterisks. Isolation and identity of fungal cultures
standard electrophoretic conditions suggested by themanufacturer.
Surface-sterilized sections from the leading edges of
A 50–350 bp DNA length standard (LI-COR. Inc.,
lesions were placed in water agar. Small tufts of
Nebraska, USA) was used as molecular weight marker
mycelia that emerged around lesion borders were trans-
for allele size calling. DNA from the isolate C-74 was
ferred to potato dextrose agar and finally multiple-
included as a reference marker in all gels, and all frag-
conidial isolates were purified. Single-conidial isolates
ments were scored against the two standards to ensure
were dissected directly from seeding spores from the
accurate designation of bands to their respective loci
infected leaf tissue, under a stereo-microscope. Cri-
over all genotypes. Isolate genotypes were screened
teria for morphological identification of Alternaria
for presence or absence (1–0) of all AFLP frag-
isolates were based on Ellis (1971). Alternaria spp.
ments between 50 and 350 bp using the electrophoresis
isolates from international sources were received as
analysis software RFLPscan v2.1 (Scananalitics-CSPI,
purified cultures. All isolates were stored in the culture
Billerica, MA, USA). Lanes and bands were tracked
manually and a binary data matrix was generateddescribing the presence or absence of bands at allscored loci for the three primer pairs. Growth of mycelium and DNA extraction
Mycelia were produced in 250 ml Erlenmeyer flasksfilled with 40 ml of potato dextrose broth. Flasks were
Similarity matrices (DICE, JACCARD and SIMPLE)
inoculated with tufts of mycelium and incubated in the
produced using the WINDIST software (I.W. Yap,
dark at 27 ±0.5 ◦C for 4 days in still culture. Mycelium
University of Washington, USA) were used to clus-
was harvested, vacuum-filtered and freezed at −80 ◦C
ter the data with three algorithms (Single linkage,
until use. DNA from each isolate was extracted from
UPGMA and Complete linkage) using the SAHN mod-
0.3–0.6 g of freeze-dried mycelium based on the extrac-
ule of NTSYSpc (v2.01, Exeter Software, Setauket,
tion method of Sambrook et al. (1989). The DNA
NY, USA). Cophenetic values were calculated using the
samples were re-extracted once to reduce discoloura-
MXCOMP module. Dendrograms were constructed
tion and RNA was degraded by addition of 1 µl RNAse
with the SAHN module using the similarity matrices
(10 mg ml−1). After quantification using a fluorometer,
from the three primer combinations considering all
samples were diluted to 50 ng µl−1 in TE buffer and
542 polymorphic fragments, and alternatively with a
reduced data set of 317 fragments in which loci forwhich all isolates were identical were pooled.
Due to the small genome of A. solani, a modified AFLPtechnique was applied to ensure amplification of suffi-
Population analysis was performed with the soft-
cient polymorphic bands. Genomic DNA from each
ware AMOVA (v1.5, Excoffier, University of Geneva,
isolate was restricted with EcoRI and MseI and ligated
Switzerland) using the reduced data set of 317 frag-
to PCR adapters using the AFLP kit of Gibco, BRL
ments. AMOVA input files were prepared using
(Gaithersburg, MD, USA). A two-step PCR procedure
AMOVA-PREP v1.01 (Mark P. Miller, University of
was adopted for selective amplification. In the first
Northern Arizona, USA). Pair-wise comparisons of the
step (pre-amplification), primers with a single selective
variation between populations were analysed by calcu-
nucleotide extension (A) were used for both the MseI
st values, which represent the proportion of the
and the EcoRI fragment ends. In the second ampli-
total variance that is partitioned between populations
fication, MseI and IRD800-labelled EcoRI primers
and measure the genetic differentiation of subpop-
with one and two selective nucleotides, respectively,
were used in combination (M-A and E-AT/-AC/-AG).
by non-parametric permutation procedures (Excoffier
AFLP products were separated in polyacrylamide gels
et al., 1992). The robustness of the phenogram was
with 1 × TBE buffer on a LI-COR 4200 fluorescent
determined by bootstrap analysis of 1000 bootstrapped
DNA analyser (LI-COR Inc., Nebraska, USA) using
samples using WINBOOT (Yap and Nelson, 1996).
A 70% bootstrapping frequency was used as the lowerlimit for recognition of robust clusters.
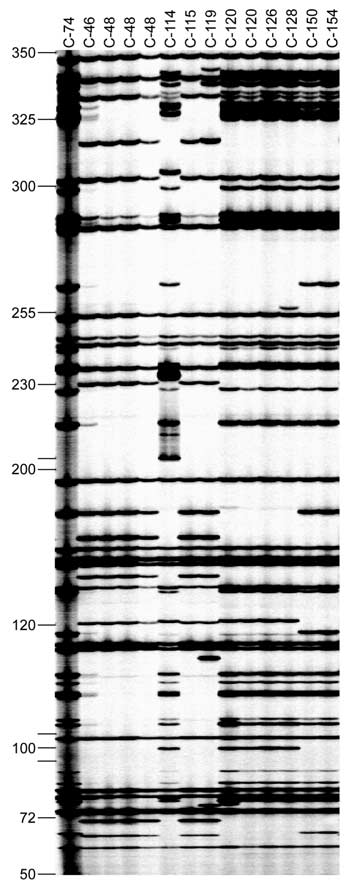
An example of typical AFLP variation is shown inFigure 1. A total of 542 fragments were observed in allisolates using the three selected primer combinations(Table 2). All primer combinations showed 100% poly-morphism for the complete set of isolates, whereas thedegree of polymorphism ranged from 89.2% to 98.6%in different isolate subsets. The primer combinationE-AT detected more fragments for all subsets of iso-lates, but was less polymorphic than E-AC and E-AGin the isolate subsets.
Dendrograms constructed using three similarity co-efficients (DICE, JACCARD and SIMPLE) and threeclustering methods (UPGMA, Single linkage and Com-plete linkage) were examined and compared to evaluatethe goodness of fit of the resulting phylogenetic treeswith respect to the reliability and stability of theinferred relationships (Table 3). High cophenetic values(ranging from 0.94 to 0.99) were found with all com-binations, both with all 542 fragments and also withthe reduced data set of 317 independent loci. In generalr > 0.9 indicates a very good fit, indicating no majorvariation among the dendrogram patterns or the threedistance calculation methods.
To assess the usefulness of AFLPs as phenetic
markers, a similarity matrix based on the SIMPLE co-efficient and UPGMA cluster method was constructedto estimate the level of relatedness among the A. solani,A. porri, Alternaria spp. and Curvularia sp. isolates. The resulting dendrogram (Figure 2) formed two mainclusters with extremely high bootstrap values that
Figure 1. Example of AFLP fingerprints in 14 isolates of
clearly distinguished the species, with the exception
Alternaria solani using the primer combination M-A/E-AT with
of the single A. porri isolate which cosegregated with
nucleotide (A) and two selective EcoRI
nucleotides (AT), respectively. Details of isolates are given in
the A. solani isolates. All A. solani isolates grouped
together in the first cluster (Cluster 1), whereas theother cluster grouped all Alternaria spp. together withthe A. alternata and Curvularia sp. isolates (Cluster 2).
being assigned to subcluster 1.1 and the majority of
Within Cluster 1, two distinct branches were con-
the potato isolates grouping in subcluster 1.2, with a
sistently formed in 100% of the 1000 bootstrapped
similarity coefficient of around 0.76 between the two
trees. These subclusters revealed a general cluster-
host-associated clusters. Subcluster 1.1 represents a
ing by host, with most A. solani isolates from tomato
large branch containing 87 genetically similar isolates
Table 2. AFLP fragments amplified in isolates of A. solani and related species with different EcoRI primerselective nucleotides. In all cases the MseI primer included one selective nucleotide (A)
a Total number of fragments observed. b Percentage of total fragments in a given species that were polymorphic. Table 3. Comparison of cophenetic values obtained from three similarity coefficients andthree clustering used for analysing the AFLP data
a Cophenetic values considering all 542 detected fragments. b Cophenetic values considering only the 317 unequal fragments.
(similarity coefficient >0.86), including 74 isolates
hand, not all isolates from Brazil clustered together,
from tomato and 12 from potato. Statistical support
although they all came from the Lavras region. Three
for minor subgroups containing isolates of different
Turkish isolates (T-4, T-5 and T-6) from tomato formed
geographic origin was poor (bootstrap values <60%),
a separate subgroup in subcluster 1.2 with a higher
however the A. solani isolates from more exotic sources
genetic distance to the other isolates, albeit with a
(China, Greece, Turkey) generally tended to group
lower statistical support of only 63%.
separately from the Cuban and Brazilian A. solani
Cluster 2 is comprised of nine isolates from tomato,
isolates, with some exceptions. The Cuban isolate
from different substrates and with different geograph-
C-240 appears to be genetically distinct from the rest
ical origins. These included the airborne isolate of
of subcluster 1.1, as it grouped separately from all
A. alternata, six isolates of Alternaria spp. with small
other isolates with 93% confidence. The isolates form-
catenulated spores and the Curvularia sp. isolate. The
ing subcluster 1.1 remained almost unchanged when
dendrogram revealed great genetic dissimilarity within
the phenogram was generated using other similarity
this cluster, with high statistical support for the sub-
indices. The A. porri isolate from onion also clustered
groups. The three isolates from Russia were almost
identical, as were the two isolates from Canada. The
The other major branch within Cluster 1 (subclus-
Curvularia and A. alternata isolates were genetically
ter 1.2) comprises 26 A. solani isolates, including 21
from potato and 5 from tomato. Of all 34 isolatesoriginating from potato, 65% grouped within thissubcluster (Figure 2). Isolates from different origins
within the USA, Greece and Turkey clustered sepa-rately to isolates originating from Cuba and Brazil,
AMOVA was used to estimate and partition the total
with a similarity coefficient of 0.80. All isolates from
genetic variance into within- and between-subgroup
the USA clustered together in a single subgroup within
subcluster 1.2 at confidence level of 100%, although
were made separately between different populations,
they originated from different states. On the other
considering the species, hosts and countries of origin. Figure 2. Phenogram showing genetic similarity among 112 isolates of Alternaria solani, seven isolates of catenulated Alternaria spp. and one isolate each of A. porri, A. alternata and Curvularia sp., revealed by UPGMA cluster analysis based on AFLP genetic fingerprintsobtained with three primer combinations. Numbers shown above branches represent percentage confidence limits obtained by bootstrapanalysis with 1000 bootstrap repetitions. Confidence limits below 70% are not shown. The isolates are separated into two highly significantclusters with a similarity coefficient of around 0.60, and the larger cluster is further divided into two subclusters with a similarity coefficientof less than 0.80.
Of the 112 A. solani isolates, 105 formed different hap-
were found between the population representing USA
lotypes with the three primer combinations tested, indi-
isolates and seven of the other eight populations anal-
cating a great genetic diversity. Considering A. solani
ysed. The Cuban populations from tomato and potato
(together with the A. porri isolate), all catenulated
showed a moderate but significant genetic difference;
Alternaria isolates and Curvularia sp. as separated
the same was observed between Turkish isolates,
populations, most of the total variability resided among
whereas between the isolates from Greece the differ-
ences were quite large ( st = 0.9591, P < 0.0001).
54.15% of the total variability) and the rest within pop-
A pair-wise comparison between the domestic Cuban
ulations. After re-configuring the data to represent six
and foreign tomato and potato isolates, respectively,
different populations consisting of A. solani isolates
revealed highly significant (P < 0.0001) genetic dif-
from Cuba, Brazil, USA, Greece, Turkey and China,
ferences between the Cuban isolate populations and the
46.35% of the total genetic variability was found to
reside between countries ( st = 0.4635). Considering
Genetic variability among the Cuban provincial
A. solani isolates from tomato and potato as two differ-
populations ( st = 0.1289) was considerably lower
ent populations, the genetic distance among them was
than among populations from different countries
0.330 with high statistical significance (P < 0.0001).
( st = 0.4635). This was consistent with the low
Table 4 shows genetic differences explained by
geographical origins of populations from tomato
Table 5. Genetic distances among the Cuban and foreign isolates
and potato, respectively. The greatest genetic dis-
of A. solani from tomato and potato, respectively, calculated using
tances among populations from tomato were observed
between the isolates from Greece and China ( st =0.6923, P < 0.0001), and between the populations
from Cuba and those from Greece and Turkey ( st =
st = 0.4560, respectively; P < 0.0001).
Among potato populations, the greatest genetic dis-
tances were found between the USA population and
those from Cuba and Brazil. All genetic distances
within this group showed highly significant differ-
ences, with the exception of the populations from
Numbers below the diagonal are measures of inter-population
pair-wise populations comparison between different
genetic distance ( st). Numbers above the diagonal are the prob-
hosts revealed a great genetic differentiation. The
st value will be greater than the observed
value, and represent the significance of the observed
Table 4. Genetic distances among nine populations representing different geographic origins and hosts of 112 A. solani isolates,calculated using three AFLP primer combinations
Numbers below the diagonal are measures of inter-population genetic distance ( st). Numbers above the diagonal are theprobabilities that a random
st value will be greater than the observed value, and represent the significance of the observed
Table 6. Genetic distances among eight populations comprising 80 A. solani isolates from different provinces of Cuba, measuredusing three AFLP primer combinations
Numbers below the diagonal are measures of inter-population genetic distance ( st). Numbers above the diagonal are the proba-bilities that a random
st values will be greater than the observed value, and represent the significance of the observed
bootstrap confidence levels for subcluster 1.1 in the
of A. solani of 0.28, whereas Weir et al. (1998) revealed
UPGMA cluster analysis (Figure 2). Results of a
a distance of 0.23. A similar level of diversity was
pair-wise comparison between the eight A. solani pop-
observed in the present study, with a minimal genetic
ulations from different Cuban provinces are shown in
similarity coefficient of around 0.76 at polymorphic
Table 6. The greatest genetic distance among popula-
AFLP loci for A. solani isolates from Cuban and
tions was observed between the samples from Pinar
del Rio and Granma ( st = 0.5113; P < 0.0001),
Phenetic and population analysis of the AFLP fin-
however the Pinar del Rio sample consists of only one
gerprint data revealed an association between genetic
isolate. Considering only the significant distances at the
background and host origin. A highly significant
P = 0.01 level, Granma is the most distinct population
genetic differentiation ( st > 0.1544; P < 0.0001)
from the others with six significant distances ( st =
was revealed among Cuban, Greek and Turkish iso-
0.1478–0.5113; P < 0.0001), followed by Camag¨uey
lates from tomato and potato (Tables 4 and 5). Evidence
( st = 0.1007–0.2201; P < 0.0001) and Holgu´ın
for host specialization of A. solani was apparent from
( st = 0.0757–0.1478; P < 0.0001) with four sig-
the phenogram of 112 isolates revealed by UPGMA
nificant distances each. Tunas showed three significant
cluster analysis. Sixty-two percent of the isolates orig-
distances ( st = 0.0757–0.1517; P < 0.0001) and the
inating from potato and 86.7% from tomato clustered
clearly in two separate branches in the dendrogram witha maximum bootstrapping confidence level (100%). AMOVA analysis with no consideration of geographic
Discussion
origin indicated a considerable genetic differentiation( st = 0.289) with high statistical significance (P <Alternaria solani is distributed all over the world where
0.0001) between the isolates representing populations
tomato and potato are cultivated (Rotem, 1994). In this
from tomato and potato. Franco et al. (2001) considered
paper, we describe a large-scale survey of genetic vari-
st values between 0.15 and 0.25 and above 0.25 to rep-
ability in 122 isolates of A. solani and related species,
resent moderately high and high genetic differentiation
from tomato and potato hosts, using AFLP genetic fin-
gerprinting. A modified AFLP protocol was applied,
To date, evidence of pathogenic specialization
optimized for the small size of the Alternaria genome
of A. solani on tomato and potato has not been
by the use of PCR primers with only one and two
reported. However, the AFLP evidence presented
selective nucleotides, respectively.
here is consistent with preliminary observations of
The AFLP data corroborated the high genetic vari-
pathogenicity in some of the A. solani isolates used in
ability reported previously for A. solani populations
this study. Isolates from potato were less aggressive
using isozymes and RAPD markers (Petrunak and
on different tomato genotypes than the tomato isolates
Christ, 1992; Weir et al., 1998). Petrunak and Christ
(data not presented). Information on the specialization
(1992) estimated a genetic distance within a population
of pathogen populations and detailed monitoring of
their pathogenicity are essential for the development of
reproduction is essential to the long-term viability of
effective disease control strategies.
a species, because it provides phenotypic variation on
In regard to pathogenic specialization, Simmons
which selection may act (Gordon and Martyn, 1997).
With the exception of a single report (Esquivel, 1984)
tomatophila, within the well-known A. solani, report-
the sexual stage in A. solani has been reported neither in
ing that A. tomatophila is ‘the common and widely
nature nor in vitro (Simmons, 1992). The genetic vari-
distributed incitant of early blight of tomato’. All iso-
ability may therefore arise from a parasexual cycle, as is
lates but one from potato examined in that study were
the case in other imperfect fungi. However, evidence of
A. solani. Host specialization in A. solani was indi-
natural parasexualism has not been obtained to date in
cated by the general separation of tomato and potato
A. solani, or in the well-known A. alternata pathogens
isolates into subclusters 1.1 and 1.2 with a genetic
(Akamatsu et al., 1999; Salamiah et al., 2001). Het-
similarity coefficient of 0.7 (Figure 2). However, some
erokaryons (Tsuge et al., 1987) and stable fusants have
isolates were not consistent with a general host spe-
been purified (Salamiah et al., 2001).
cialization hypothesis. Nine potato isolates from Cuba,
Because of the low number of catenulated Alternaria
two from Turkey and two from Brazil were grouped
spp. isolates studied it is difficult to make conclusions
among tomato isolates in subcluster 1.1, whereas one
regarding their genetic similarity to other isolates.
Cuban and three Turkish tomato isolates grouped
However, our results are consistent with earlier studies
within the potato isolates in subcluster 1.2. We have
that found high variability within catenulated isolates
therefore classified all potato and tomato isolates
(Morris et al., 2000; Roberts et al., 2000; Andersen
in A. solani, according to Ellis (1971), until detailed
et al., 2001). Moreover, in our experiment the catenu-
microscopic and cultural examination according to
lated isolates showed a low genetic similarity (0.61)
in relation to the A. solani isolates. In other studies
The results of the present study revealed an influ-
A. solani and A. alternata were also clearly distin-
ence of geographic origin on genetic variability among
guishable from each other (Petrunak and Christ, 1992;
the populations of A. solani. Isolates from the USA,
Weir et al., 1998). Three Alternaria species pathogenic
Brazil, Cuba and Turkey were genetically distant from
to crucifers with small and large spores were also
distinguishable using RAPD analysis (Sharma and
tions within Cuba showed little significant genetic
diversity (Table 6). This lack of significant geographic
This study demonstrates the suitability of the AFLP
differentiation among regional Alternaria spp. pop-
technique for detailed analysis of genetic variation
ulations suggests (1) widespread dispersal of fungal
in A. solani and related pathogens. Cluster analysis
spores and (2) weak host selection pressure within
indicated a level of host specificity within A. solani.
and between populations (Sharma and Tewari, 1998;
This may have important implications for effective
early blight disease management in tomato and potato.
A high degree of morphological and DNA similar-
It appears that the use of reproducible, highly poly-
ity among A. solani and A. porri has been observed
morphic AFLP markers has the potential to play a
(Neergaard, 1945; Pryor and Gilbertson, 2000).
major role in the accurate taxonomic identification of
The members of the porri species-group (A. porri,
A. solani, Alternaria dauci, Alternaria macrospora andAlternaria crassa) exhibit a high degree of rDNA sim-ilarity, with no differences or only minor variation in
Acknowledgements
mitochondrial small subunit or nuclear internal tran-scribed spacer sequences. Based on this evidence, an
We are grateful to Drs. E. Demirci and S. Benlioglu
extremely close relationship among species in this
(Turkey), I. Vloutoglou and Y. Manetas (Greece), Y.H.
group has been suggested (Pryor and Gilbertson, 2000),
Tong (China), C. Wharam (USA), T.A. Evstigneeva
and this was clearly reflected by the inability of the
(Russia), M. Tu (Canada), P. Nurmberg (Brazil),
AFLP analysis to distinguish the A. porri isolate from
M.A. Dita and R. Casta˜neda (Cuba) for providing iso-
A. solani in the present study.
lates used in this study. We also thank Petra Theuer
The origin of the variability present in A. solani
for technical assistance. SPM was supported by a grant
is unknown. It is commonly assumed that sexual
from DAAD (German Academic Exchange Service). References
Peever TL, Olsen L, Iba˜nes A and Timmer LW (1999)
Genetic differentiation and host specificity among popula-tions of Alternaria spp. causing brown spot of Grapefruit and
Adachi Y, Watanabe H and Tsuge T (1996) Relationships
Tangarina × Grapefruit hybrids in Florida. Phytopathology 90:
between genetic polymorphisms and fungicide resistance
within Alternaria alternata. Phytopathology 86: 1248–1254.
Pei MH and Ruiz C (2000) AFLP evidence of the distinctive
Akamatsu H, Taga M, Kodama M, Johnson M, Otani R and
patterns of life-cycle in two forms of Melampsora rust on Salix
Kohmoto K (1999) Molecular karyotypes for Alternaria plant
viminalis. Mycological Research 104: 937–942.
pathogens known to produce host-specific toxins. Current
Petrunak DM and Christ BJ (1992) Isozyme variability in
Alternaria solani and Alternaria alternata. Phytopathology 82:
Andersen B, Kroger E and Roberts RG (2001) Chemical and
morphological segregation of Alternaria alternata, A. gaisen
Pryor BM and Gilbertson RL (2000) Molecular phylogenetic rela-
and A. longipes. Mycological Research 105: 291–299.
tionships amongst Alternaria species and related fungi based
Aradhya MK, Chan HM and Parfitt DE (2001) Genetic variabil-
upon analysis of nuclear ITS and mt SSU rDNA sequences.
ity in the pistachio late blight fungus, Alternaria alternata.
Mycological Research 104: 1312–1321.
Mycological Research 105: 300–306.
Roberts RG, Reymond ST and Andersen B (2000) RAPD frag-
Bock CH, Thrall PH, Brubaker CL and Burdon JJ (2002) Detec-
ment pattern analysis and morphological segregation of small-
tion of genetic variation in Alternaria brassicicola using AFLP
spored Alternaria species and species groups. Mycological
fingerprinting. Mycological Research 106: 428–434.
Bonde R (1929) Physiological strains of Alternaria solani.
Rotem J (1966) Variability in Alternaria porri f.sp. solani. Israel
Bruner SC (1918) La pudrici´on del tomate y modo de evitarla.
Rotem J (1994) The Genus Alternaria: Biology, Epidemiology
Revista de Agricultura, Comercio y Trabajo 1: 300–303. and Pathogenicity. APS Press, St. Paul, Minnesota.
Ellis JB and Martin GB (1882) New species of North Amer-
Salamiah A, Akamatsu H, Fukumasa-Nakai H, Otani H and
ican fungi: Macrosporium solani. American Naturalist 16:
Kodama M (2001) Construction and genetic analysis of hybrid
strains between apple and tomato pathotypes of Alternaria
Ellis MB (1971) Dematiaceous Hyphomycetes. Commonwealth
alternata by protoplast fusion. Journal of General Plant Pathol-
Mycological Institute, FAO, Kew, Surrey.
Esquivel EA (1984) Pleospora solani sp-nov, telemorphosis of
Sambrook J, Fritsh EF and Maniatis T (1989) MolecularAlternaria solani (Ell and Mart) Jones and Grout. Phytopathol-
Cloning: A Laboratory Manual. Cold Spring Laboratory Press,
Excoffier L, Smouse PE and Quattro JM (1992) Analysis of
Sharma TRT and Tewari JP (1998) RAPD analysis of three
molecular variance inferred from metric distance among DNA
Alternaria species pathogenic to crucifers. Mycological
haplotypes: Application to human mitochondrial DNA restric-
Simmons EG (1992) Alternaria taxonomy: Currents status, view-
Franco J, Crossa J, Ribaut JM, Betran J, Warburton ML and
point, challenge. In: Chelkowski J and Visconti A (eds)
Khairallah M (2001) A method for combining molecular mark-
Alternaria: Biology, Plant Diseases and Metabolites. Elsevier
ers and phenotypic attributes for classifying plant genotypes.
Science Publishers, Amsterdam, pp. 1–35.
Theoretical and Applied Genetics 103: 944–952.
Simmons EG (2000) Alternaria themes and variations (244–286),
Gordon TR and Martyn RD (1997) The evolutionary biology of
species on solanaceae. Mycotaxon, LXXV: 1–115. Fusarium oxysporum. Annual Reviews of Phytopathology 35:
Stancheva Y (1989) Investigations of the phytotoxic activity
of culture filtrates of different isolates of Alternaria solani.
Izquierdo F (1981) Evidencia de la existencia de razas fisiol´ogicas
en Alternaria solani. Revista de Ciencias Biol´ogicas 12:
Tsuge T, Hayashi N and Nishimura S (1987) Selection of
auxotrophic mutants and heterokaryosis in Alternaria alter-
Mart´ınez B, Bernal A, P´erez S and Munis Y (2002) Variabilidad
nata. Annals of the Phytopathological Society of Japan 53:
patog´enica de aislamientos en Alternaria solani Sor. Revista
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes
McDonald BA (1997) The population genetics of fungi: Tools
M, Frijters A, Pot J, Peleman J and Kuiper M (1995) AFLP:
and techniques. Phytopathology 87: 448–453.
A new technique for DNA fingerprinting. Nucleic Acids and
Morris PF, Connolly MS and St Clair DA (2000) Genetic diver-
sity of Alternaria alternata isolated from tomato in California
Weir TL, Huff DR, Christ BJ and Romaine CP (1998) RAPD-PCR
assessed using RAPDs. Mycological Research 104: 286–292.
analysis of genetic variation among isolates of Alternaria solani
and Alternaria alternata from potato and tomato. Mycologia
Pathogenit¨at von Alternaria porri (Ell.) Saw. f. sp. solani E.
et M. Journal of Phytopathology 41: 317–352.
Yap IV and Nelson RJ (1996) WinBoot: A program for performing
Neergaard P (1945) Danish Species of Alternaria and Stem-
bootstrap analysis of binary data to determine the confidence
phylium. Taxonomy, Parasitism, Economical Significance.
limits of UPGMA-based dendrograms. IRRI Discussion Paper
Humphrey Millford, Oxford University Press, London.
Series No. 14, International Rice Research Institute, Manila.
Double Anaerobic Coverage: What is the role in clinical practice? BACKGROUND Anaerobic pathogens are normal flora of the oral cavity and the gastrointestinal tract. While oral anaerobic flora are mostly gram-positive organisms such as Peptococcus and Peptostreptococcus spp., the principal anaerobic intestinal flora are gram-negative bacil i such as Bacteroides fragilis , Prevotel a m
The names of the Holocaust victims that appear on this list were taken from Pages of Testimony submitted to Yad Vashem First name Family name Place of birth Date of birth Age Place of residence Place of death Date of death www.yadvashem.org The names of the Holocaust victims that appear on this list were taken from Pages of Testimony submitted to Yad Vashem First name F
 A 70% bootstrapping frequency was used as the lowerlimit for recognition of robust clusters.
A 70% bootstrapping frequency was used as the lowerlimit for recognition of robust clusters.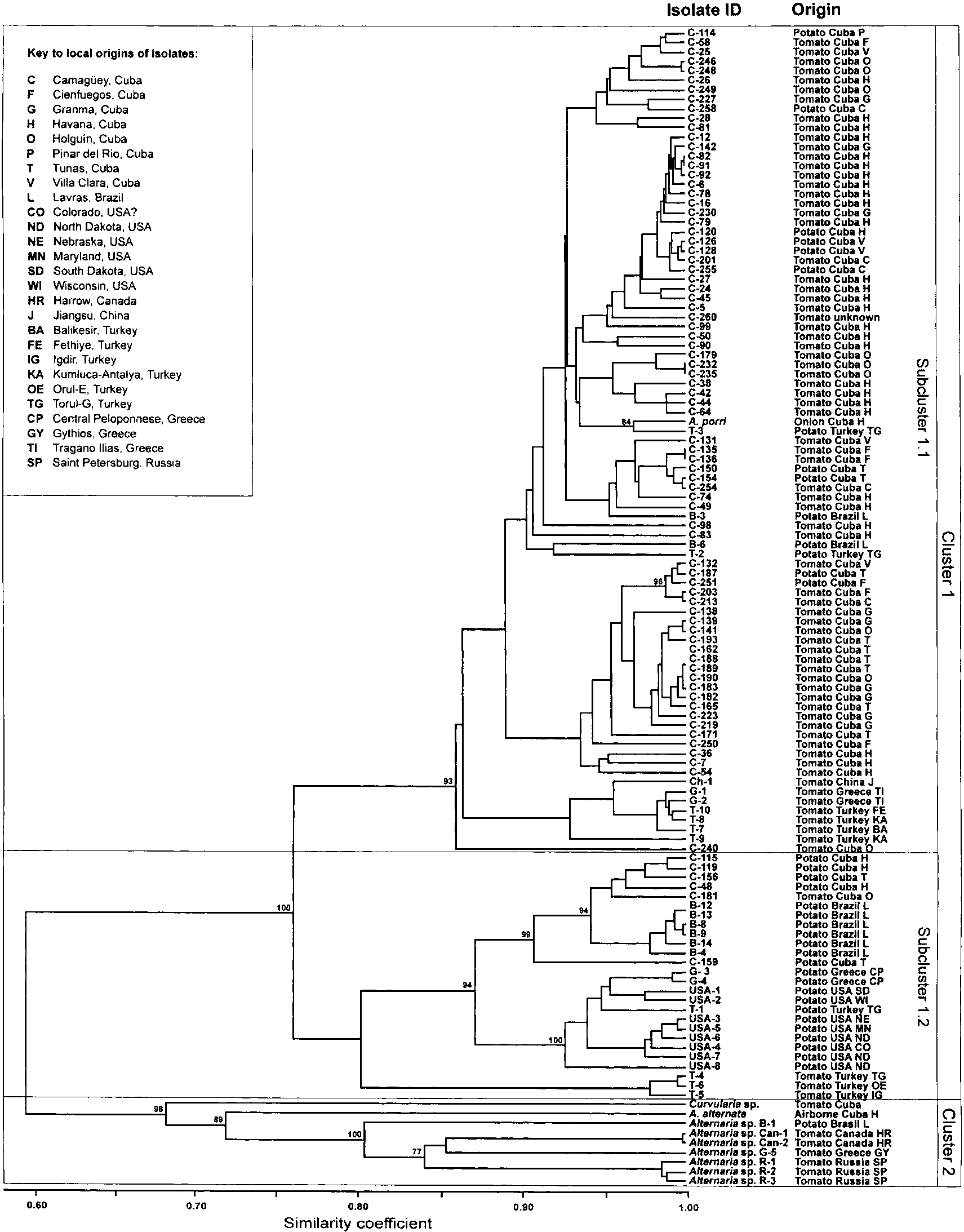 Figure 2. Phenogram showing genetic similarity among 112 isolates of Alternaria solani, seven isolates of catenulated Alternaria spp.
Figure 2. Phenogram showing genetic similarity among 112 isolates of Alternaria solani, seven isolates of catenulated Alternaria spp.