Thank you, it's cool. Used for the first time because only then was the right drug. Very revealing detail: ordered Express shipping to get the order within 3 hours because didn't know how to get the order faster https://africarx.co.za/buy-levitra-south-africa.html The drug is authentic exactly. Consistent with the stated prices the Staff is knowledgeable.
Uasr18.3.-17972 295.299
Arid Land Research and Management 18: 295–299, 2004Copyright # Taylor & FrancisISSN: 1532-4982 print/1532-4990 onlineDOI: 10.1080/15324980490451483
Research Note: Spatial Variation of AM Fungal
Spore Numbers under Canopies of Acacia raddiana
Faculty of Life SciencesNorthwest Science and TechnologyUniversity of Agriculture and ForestryShaanxi, P.R. China and Faculty of Life SciencesBar-Ilan University, Ramat-Gan, Israel
Faculty of Life SciencesBar-Ilan UniversityRamat-Gan, Israel
The spatial variation of arbuscular mycorrhizal fungal spores under the canopy ofAcacia raddiana was studied in a desert system. Soils samples from the base of thestem, its canopy radius, and outside its canopy at four stations in the Negev desertwere collected from a 0 to 50 cm depth at sections of 10 cm each. The mean sporedensity was found to fluctuate between 265 100 gÀ1 to 105 100 gÀ1 in fluvisols andcalcaric fluvisols, respectively. Our results suggest that spore density and sporedistribution were found to be directly correlated with soil type, elucidating theimportance of soil physical composition on AM fungi distribution in desert soilecosystems.
arid ecosystem, Negev Desert, desert soils, fungal distribution
Arbuscular mycorrhizal fungal (AMF) associations are ubiquitous in desertecosystems and may play an important role in plant establishment and growth bybridging between plant and soil (Allen, 1983; Dhillion & Zak, 1993; Skujin¸sˇ & Allen,1986). The abundance of arbuscular mycorrhizal fungi and their spores are depen-dent on type and physical characteristics of soil (Bethlenfalvay et al., 1988;Ortega-Larrocea et al., 2001).
Bever et al. (1996) and Sanders & Fitter (1992) have postulated that the host
plant may play a significant role in every phase of the life history of AM fungi (sporegermination, hyphal development and colonization, and sporulation) by regulatingcarbon allocation to roots, producing secondary metabolites, or changing soilenvironmental conditions. Therefore, investigating the spatial distribution andrichness of AM fungal spores related to host plants in the Negev Desert might be
Received 11 October 2003; accepted 20 January 2004. Address correspondence to Prof. Y. Steinberger, Faculty of Life Sciences, Bar-Ilan
University, Ramat-Gan 52900, Israel. E-mail: steinby@mail.biu.ac.il
helpful for understanding the ecological role of AM fungi in desert ecosystems andthe relationship between AM fungal distribution and the growth of desert plants (Heet al., 2002a, 2002b).
We hypothesized that sampling location soil type and depth will strongly reflect
the abundance and distribution of AM fungal spores in a desert ecosystem. The aimof this study was to determine the influence of soil type on the abundance anddistribution of AM fungal spores in the A. raddiana rhizosphere in a desert soilecosystem.
Soil samples were collected at four sites along a soil type gradient running from
the northern Negev Desert 25 km east of Dimona (30058oN; 35020oE), with lithosoltype soil, mean annual temperature of 20oC and mean annual rainfall of 150 mm,south toward the Dead Sea, in the Arava Valley-Ein Ofarin (30045oN; 35018oE), withcalcaric fluvisol type soil, mean yearly temperature ranging between 22 and 24oC andmean rainfall of 42 mm, southwest toward the Faran Desert (30012oN; 34055oE), withfluvisol type soil, mean annual temperature range between 20 and 22oC, meanannual rainfall of approximately 50 mm, and to the north near Sede Boker, RamatBoker (30048oN; 34045oE), with yermosol type soil, mean annual temperature of18oC and mean annual rainfall of 97 mm (Dan et al., 1972, 1977). All four sitesrepresent arid conditions with a common feature of low and unpredictable rainfall,desert climate, mild, rainy winters and hot, dry summers.
Acacia raddiana Savi is a typical summer-active shrub that grows in many wadis.
It originates from the savannahs of Africa and penetrated the Negev from the farsouth (Evenari et al., 1982). Recent studies had shown an increase in VAM colo-nization in the root area of Acacia, reflecting vigorous growth of these trees (Ishii,2000).
Soil samples at the base of the A. raddiana stem, the canopy radius (2 m), and outsidethe canopy (2 m from canopy edge) as control (interplant) were collected at the foursites in the summer of 2000. The soil samples were collected in three replicates from adepth of 50 cm at each location and were divided into sections corresponding to0–10, 10–20, 20–30, 30–40 and 40–50 cm depths. They were placed in individualplastic bags and transported to the laboratory in an insulated container and sieved(2 mm mesh size) before processing.
Twenty-five grams of soil from each replicate were used for spore extraction.
The total AM fungal spore number was determined by wet sieving (45–500 mm),sucrose density centrifugation (Ianson & Allen, 1986) and counting under astereoscopic microscope at Â40.
All data were subjected to statistical analysis of variance using the SAS model
(ANOVA, Duncan’s multiple range test and correlation coefficient) and were used toevaluate differences between separate means.
The mean spore density was found to be significantly higher (P < 0.05) at the Faransite (265 100 gÀ1 soil) than at Ramat Boker (127 100 gÀ1 soil), Dimona (162 100 gÀ1soil), and Ein Ofarin (105 100 gÀ1 soil). Similar differences were obtained betweenRamat Boker, Dimona, and Ein Ofarin. However, no significant differences(P > 0.05) were found between Ramat Boker and Ein Ofarin. Sampling locationrelative to plant stem had a significant effect on the mean spore density. These valueswere found to be significantly higher (P < 0.05) only at the base of the stem ratherthan in the canopy radius. At the 0–10 cm soil layer, the mean spore density wasfound to range between 96 100 gÀ1 soil and 268 100 gÀ1 soil, increased slightly with
FIGURE 1 Mean spore density 100 gÀ1 soil samples taken at (A) canopy base, (B)canopy radius, and (C) control, at four sites: (I) Dimona; (II) Ramat Boker; (III) EinOfarin; and (IV) Faran, under A. raddiana (
depth and reached a maximum value of 285 100 gÀ1soil (Faran site), 110 100 gÀ1 soil(Ein Ofarin) at the 20–30 cm depth and 140 100 gÀ1 soil (Ramat Boker) and 168100 gÀ1 soil (Dimona) at the 30–40 cm depth, then declined gradually with depth. A similar depth variation pattern was found in the different sampling locationsrelative to the plant stem, however, no significant differences were found betweensampling depths ( Figure 1).
Our results demonstrate that the mean spore density related to the canopy ofA. raddiana in a Negev Desert ecosystem was 165 100 gÀ1 soil, and the distribution ofspore density in the soil was extremely variable. Sampling location had a significanteffect on spore density. The mean spore density at the Faran site (265 100 gÀ1 soil)
was significantly higher than in Ramat Boker (127 100 gÀ1 soil) and Ein Ofarin(105 100 gÀ1 soil). The highest spore densities under the canopy of A. raddiana in thefour sampling stations were found at different soil depths.
This result may be closely related to soil type and not to environmental condi-
tions, which were found to be similar in soil moisture and organic matter content(Pen-Mouratov et al., 2003). At the Faran site, soil conditions were favorable forplant inhabitance and fungal reproduction, and vice versa in the other three stationswhere they were less auspicious (soil layers contain very little soil and numeroussmall stones).
The microenvironment of a relatively small gap surrounded by the tested shrub
may not be greatly affected, and the roots from the bordering shrub may invade thegap. Thus, there may be no significant difference between small gaps and under thecanopy.
Our results suggest that spore density and spore distribution were found to be
directly correlated with the soil type, elucidating the importance of soil physicalcomposition on AM fungal distribution in desert soil ecosystems.
Allen, M. F. 1983. Formation of vesicular-mycorrhizae in Atriplex gardneri (Chenopodiaceae):
Seasonal response in a cold desert. Mycologia 75:773–776.
Bethlenfalvay, G. J., M. S. Brown, and R. S. Ames. 1988. Effects of drought on host and
endophyte development in mycorrhizal soybeans in relation to water-use and phosphate-uptake. Physiologia Plantarum 72:565–571.
Bever, J. D., J. B. Morton, J. Antonovics, and P. A. Schultz. 1996. Host-dependent spor-
ulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. Journalof Ecology 84:71–82.
Dan, J., D. H. Yaalon, H. Koyumdji, and Z. Raz. 1972. The soil association map of Israel
(1:1,000,000) Israel Journal of Earth Sciences 21:29–49.
Dan, J., D. H. Yaalon, H. Koyumdji, and Z. Raz. 1977. The soils of Israel (with 1:500,000 soil
map of Israel). Bulletin 168 of the Volvani Institute of Agricultural Research, Beit Dagan,Israel (in Hebrew).
Dhillion, S. S. and J. C. Zak. 1993. Microbial dynamics in arid ecosystems - Desertification
and the potential role of mycorrhizas. Revista Chilena de Historia Natural 66:253–270.
Evenari, M. E., L. Shanan, and W. Tadmore. 1982. The Negev:The challenge of a desert.
Harvard University Press, Cambridge, Massachusetts, USA.
He, X., S. Mouratov, and Y. Steinberger. 2002a. Spatial distribution and colonization of
arbuscular mycorrhizal fungi under the canopies of desert halophytes. Arid Land Researchand Management 16:149–160.
He, X., S. Mouratov, and Y. Steinberger. 2002b. Temporal and spatial dynamics of vesicular-
arbuscular mycorrhizal fungi under the canopy of Zygophyllum dumosum Boiss. in theNegev Desert. Journal of Arid Environments 52:379–387.
Ianson, D. C. and M. F. Allen. 1986. The effects of soil texture on extraction of vesicular-
arbuscular mycorrhizal spores from arid soils. Mycologia 78:164–168.
Ishii, T. 2000. The utilization of mycorrhizal fungi on agroforestry systems in the semi arid
regions of Kenya. Scientific Report, Kyoto Pref. University. Human Environmental andAgriculture 52:21–37.
Ortega-Larrocea, M. P., C. Siebe, G. Becard, I. Mendez, and R. Webster. 2001. Impact of a
century of wastewater irrigation on the abundance of arbuscular mycorrhizal spores in thesoil of the Mezquital Valley of Mexico. Applied Soil Ecology 16:149–157.
Pen-Mouratov, S., X. He, and Y. Steinberger. 2003. Spatial distribution and trophic diversity
of nematode populations under Acacia raddiana along a temperature gradient in the NegevDesert ecosystem. Journal of Arid Environments (In press).
Sanders, I. R. and A. H. Fitter. 1992. Evidence for differential responses between host-fungus
combinations of vesicular-arbuscular mycorrhizas from a grassland. Mycological Research96:415–419.
Skujin¸sˇ, J. and M. F. Allen. 1986. Use of mycorrhizae for land rehabilitation. MIRCEN
Bt-maize and non-target organisms Basel, October 1999 1. SUMMARY . 4 2. INTRODUCTION . 4 3. EFFECTS OF BT-MAIZE ON NON-TARGET INSECTS - REGULATORY AND NON- REGULATORY REQUIRED STUDIES . 10 STUDIES ON THE IMPACT OF BT MAIZE TO NON-TARGET INSECTS.10 48-hour static renewal toxicity of pollen from the geneticallymodified maize to water fleas ( Daphnia magna) .11Single dose t
MATERIAL SAFETY DATA SHEET SAWYER’S PERMETHRIN TICK REPELLENT ODORLESS FORMULA CLOTHING TREATMENT ONLY DO NOT APPLY TO SKIN I. PRODUCT IDENTIFICATION UN NUMBER: 1950, Aerosols, Non-Flammable, N.O.S. Class 2 II. HAZARDOUS INGREDIENTS Inert Ingredients (Proprietary mixture of solvents, surfactants, preservatives, and propellents) *(3-phenoxyphenyl)methyl (+/-)
 Arid Land Research and Management 18: 295–299, 2004Copyright # Taylor & FrancisISSN: 1532-4982 print/1532-4990 onlineDOI: 10.1080/15324980490451483
Research Note: Spatial Variation of AM Fungal
Spore Numbers under Canopies of Acacia raddiana
Faculty of Life SciencesNorthwest Science and TechnologyUniversity of Agriculture and ForestryShaanxi, P.R. China and Faculty of Life SciencesBar-Ilan University, Ramat-Gan, Israel
Faculty of Life SciencesBar-Ilan UniversityRamat-Gan, Israel
The spatial variation of arbuscular mycorrhizal fungal spores under the canopy ofAcacia raddiana was studied in a desert system. Soils samples from the base of thestem, its canopy radius, and outside its canopy at four stations in the Negev desertwere collected from a 0 to 50 cm depth at sections of 10 cm each. The mean sporedensity was found to fluctuate between 265 100 gÀ1 to 105 100 gÀ1 in fluvisols andcalcaric fluvisols, respectively. Our results suggest that spore density and sporedistribution were found to be directly correlated with soil type, elucidating theimportance of soil physical composition on AM fungi distribution in desert soilecosystems.
Arid Land Research and Management 18: 295–299, 2004Copyright # Taylor & FrancisISSN: 1532-4982 print/1532-4990 onlineDOI: 10.1080/15324980490451483
Research Note: Spatial Variation of AM Fungal
Spore Numbers under Canopies of Acacia raddiana
Faculty of Life SciencesNorthwest Science and TechnologyUniversity of Agriculture and ForestryShaanxi, P.R. China and Faculty of Life SciencesBar-Ilan University, Ramat-Gan, Israel
Faculty of Life SciencesBar-Ilan UniversityRamat-Gan, Israel
The spatial variation of arbuscular mycorrhizal fungal spores under the canopy ofAcacia raddiana was studied in a desert system. Soils samples from the base of thestem, its canopy radius, and outside its canopy at four stations in the Negev desertwere collected from a 0 to 50 cm depth at sections of 10 cm each. The mean sporedensity was found to fluctuate between 265 100 gÀ1 to 105 100 gÀ1 in fluvisols andcalcaric fluvisols, respectively. Our results suggest that spore density and sporedistribution were found to be directly correlated with soil type, elucidating theimportance of soil physical composition on AM fungi distribution in desert soilecosystems.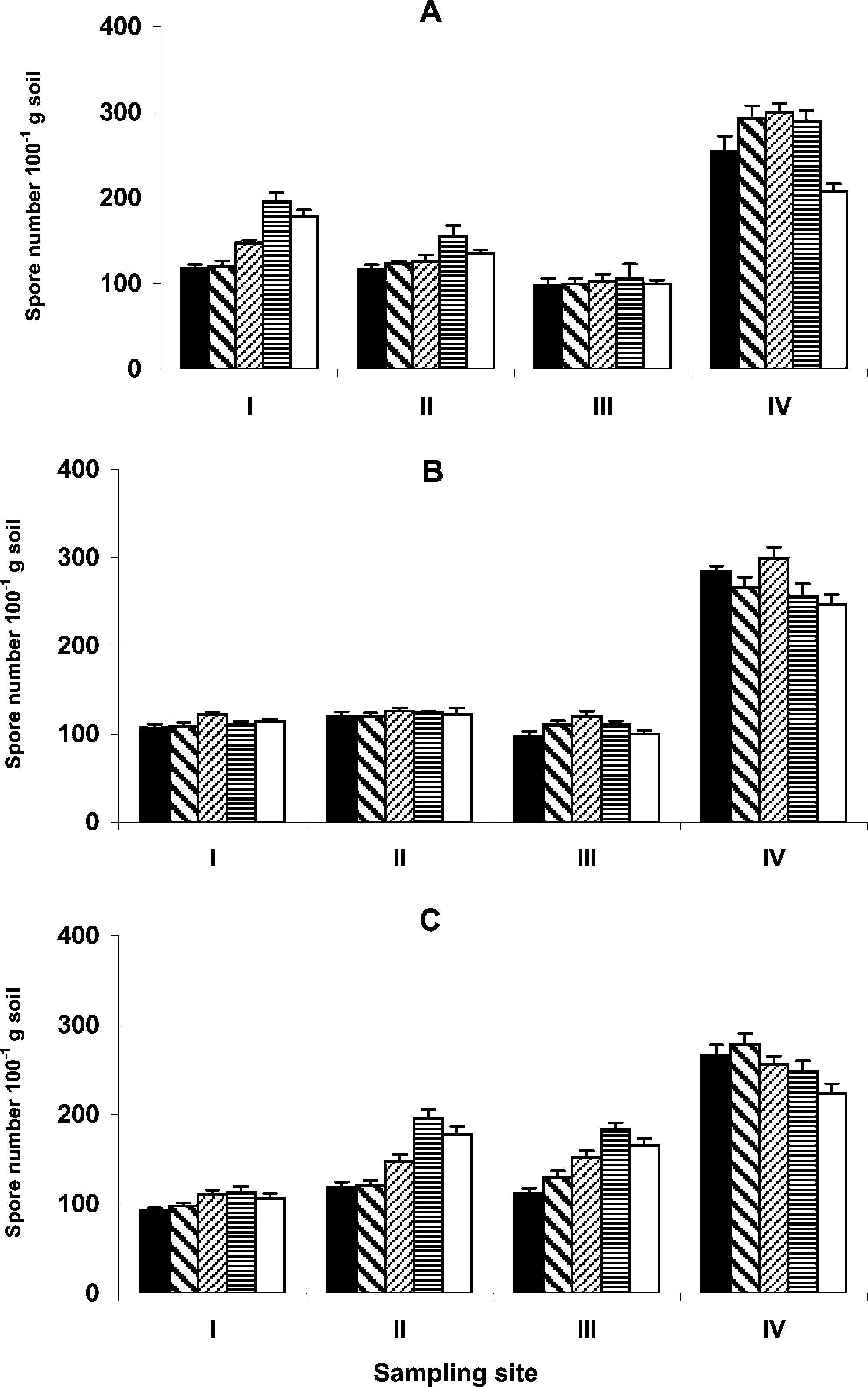




 FIGURE 1 Mean spore density 100 gÀ1 soil samples taken at (A) canopy base, (B)canopy radius, and (C) control, at four sites: (I) Dimona; (II) Ramat Boker; (III) EinOfarin; and (IV) Faran, under A. raddiana (
depth and reached a maximum value of 285 100 gÀ1soil (Faran site), 110 100 gÀ1 soil(Ein Ofarin) at the 20–30 cm depth and 140 100 gÀ1 soil (Ramat Boker) and 168100 gÀ1 soil (Dimona) at the 30–40 cm depth, then declined gradually with depth.
FIGURE 1 Mean spore density 100 gÀ1 soil samples taken at (A) canopy base, (B)canopy radius, and (C) control, at four sites: (I) Dimona; (II) Ramat Boker; (III) EinOfarin; and (IV) Faran, under A. raddiana (
depth and reached a maximum value of 285 100 gÀ1soil (Faran site), 110 100 gÀ1 soil(Ein Ofarin) at the 20–30 cm depth and 140 100 gÀ1 soil (Ramat Boker) and 168100 gÀ1 soil (Dimona) at the 30–40 cm depth, then declined gradually with depth.