Ordered, immediately called back and the same day delivered the order.Very pleased with the work. Thank you for prompt and accurate work https://africarx.co.za/buy-levitra-south-africa.html great prices, delivered on the day of the order. Pleasant managers consult by phone.
Unprecedented diversity of catalytic domains in the first four modules of the putative pederin polyketide synthase
Unprecedented Diversity of Catalytic Domains inthe First Four Modules of the Putative PederinPolyketide SynthaseJˆrn Piel,*[a] Gaiping Wen,[b] Matthias Platzer,[b] and Dequan Hui[a]
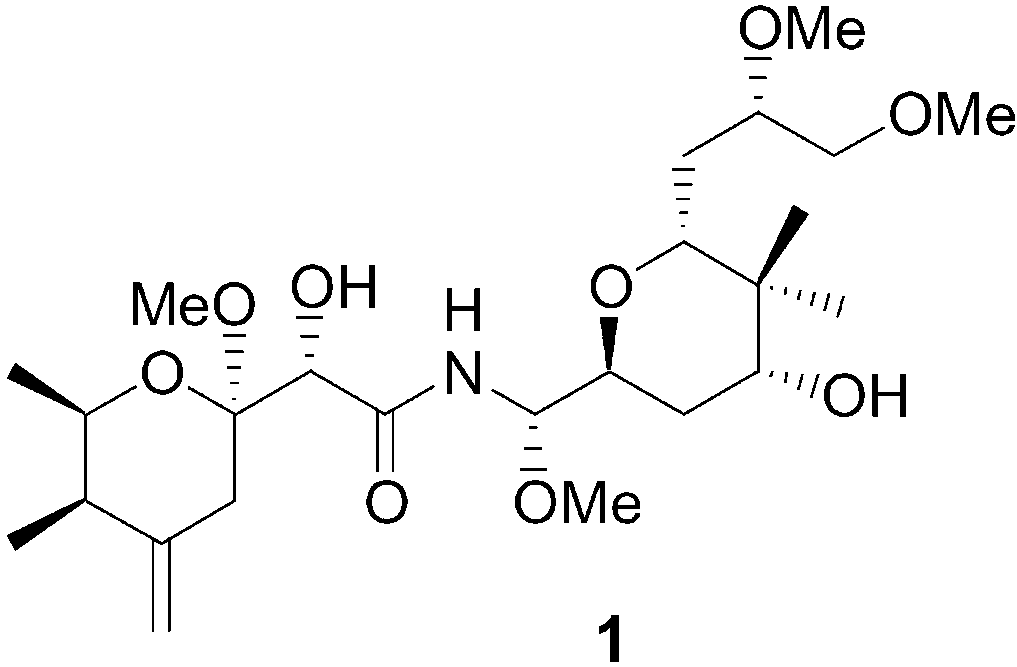
Polyketides of the pederin group are highly potent antitumor
region of the symbiont genome. It contains at least three novel
compounds found in terrestrial beetles and marine sponges.
catalytic domains that are predicted to be involved in pederin
Pederin is used by beetles of the genera Paederus and Paederidus
chain initiation and the formation of an unusual exomethylene
as a chemical defense. We have recently identified a group of
bond. The region is bordered by mobility pseudogenes; this
putative pederin biosynthesis genes and localized them to the
suggests that gene transposition led to the disjointed cluster
genome of an as yet unculturable Pseudomonas sp. symbiont, the
organization. With this work, all putative pederin genes have been
likely true pederin producer. However, this polyketide synthase
identified. Their heterologous expression in a culturable bacterium
cluster lacks several genes expected for pederin production. Here
will provide important insights into how sustainable sources of
we report an additional polyketide synthase encoded on a separate
invertebrate-derived drug candidates can be created.
It has been proposed for many years that natural productsymbiosis, that is, the biosynthesis of secondary metabolites bysymbiotic organisms, plays an important role in the chemistry ofinvertebrates.[1±4] Such suspected symbiont metabolites includemany promising drug candidates isolated from marine animals. The existence of producing bacteria could be exploited toovercome the supply problem that currently hampers animal-derived drug development and production. However, ourcurrent general inability to cultivate such symbionts representsa serious technical challenge in pinpointing the true producer. So far, the most compelling evidence for symbiotic producers
polyketide synthase (PKS) ± nonribosomal peptide synthetase
exists for the complex polyketide pederin (1), a highly potent
system, whose architecture perfectly matches a large portion ofthe pederin (1) structure.[7] Although these genes were isolatedfrom the total DNA of Paederus fuscipes beetles, they belong tothe genome of an as yet unculturable symbiotic bacterium witha very close relationship to Pseudomonas aeruginosa.[7, 8]
Bacterial type I PKSs are giant, modular enzymes harboring a
multitude of catalytic domains.[9] Each module typically consistsof at least a ketoacyl synthase (KS), an acyltransferase (AT), and
Max Planck Institute for Chemical Ecology
anticancer agent. It is found in apparently all species of the rove
Department of Bioorganic ChemistryHans-Knˆll-Strasse 8, Beutenberg Campus
beetle genera Paederus and Paederidus,[5] whose females use
pederin to chemically defend their offspring against predators.[6]
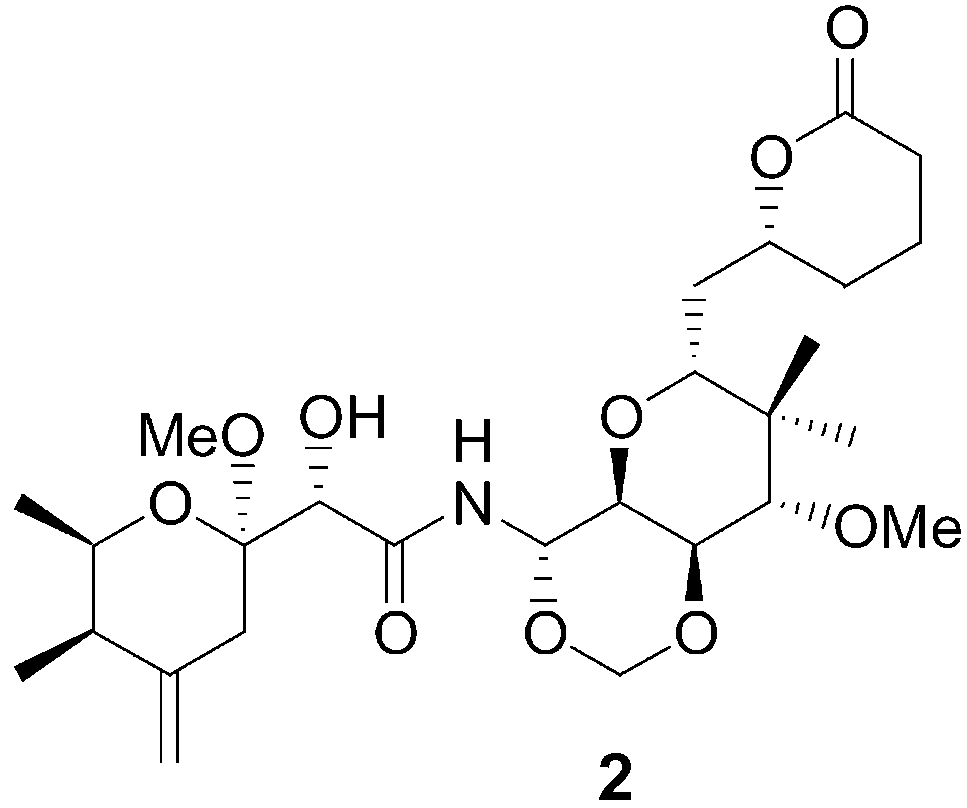
Remarkably, closely related compounds have also been isolated
from several sponges during marine antitumor and antiviral
screening programs, for example, theopederin C (2) and onna-
mide A (3) from Theonella swinhoei.[5] Recently we have reported
the ped biosynthesis gene cluster encoding a hybrid type I
¹ 2004 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
can be distinguished from those of cis-AT systems by phyloge-netic analysis.[18] Such an analysis placed the amplified productdesignated as PS3 close to the cis-AT, and PS1 and PS2 into thetrans-AT group. Since the missing part of the ped PKS shouldbelong to the latter group, we intended to isolate the gene(s)belonging to these PCR products. We therefore screened2000 clones of a cosmid library prepared from DNA of P. fuscipesbacteria with probes derived from PS1 and PS2. Unexpectedly,restriction mapping of the positive clones revealed that the twoPCR products did not originate from the same genomic region
an acyl carrier protein (ACP) domain, and attaches one polyke-
and apparently belonged to different secondary metabolites.
tide building block, usually a small malonyl-CoA derivative, to
Partial sequencing of both regions identified the PS2 system as a
the growing polyketide chain. During each of these elongation
mixed peptide synthetase ± PKS unrelated to the pederin (1)
steps, various optional additional domains present on the
structure (data not shown), while the domain arrangement of
module can introduce further structural modifications. The order
the PS1 PKS was in very good agreement with the proposed
and domain architecture of modules on the PKS enzymes usually
architecture. The cosmid pPS9D2 containing the entire PS1
strictly correspond to the order and structure of each polyketide
region, as judged from sequencing of insert ends, was therefore
building block, a phenomenon known as the colinearity rule.
This rule can be used to predict whether an isolated PKS genecluster encodes the biosynthesis of a specific metabolite. It also
permits a rational design of novel polyketides by geneticmodification of the domain architecture.[10, 11]
A map of the completely sequenced insert of cosmid pPS9D2 is
An analysis of the domain organization of the ped system
shown in Figure 1, and the deduced functions of the identified
revealed a number of surprises. The AT domains, usually found
open reading frames (ORFs) are listed in Table 1. As with the
within each module, were instead encoded by separate genes.[7]
previously sequenced ped region, several genes with remarkable
Although this was the first reported example of monofunctional
homology to Ps. aeruginosa ORFs were found in proximity to the
ATs encoded by type I PKS clusters, a number of similar systems
PKS; this indicates that the two systems are located on the same
have been discovered recently; this suggests that such enzymes
genome. These homologues comprise a cluster involved in the
might be more widespread among bacteria.[12±14] In addition,
uptake of branched-chain amino acids, a putative heat-shock
several partial gene clusters have been published, whose
gene, and a probable hemin receptor gene. In addition, the
modules lack ATs and that probably also belong to this
cosmid harbors various pseudogenes, including 11 decayed IS-
group.[15±17] Work on the leinamycin biosynthesis gene cluster
like elements (IS insert sequence). Framed by two of these ISs
revealed that these ATs acylate each module in trans,[12] and we
is a small gene cluster highly similar to PKS systems. Its three
have shown that such ™trans-AT∫ PKSs are phylogenetically
genes, designated pedI, pedJ, and pedK, do not contain frame-
distinct from regular ™cis-AT∫ systems.[18] The second deviation
shifts or stop codons and therefore seem to be intact.
from the general PKS rules was that the genes encoding the first
An inspection of the domain arrangement of PedI revealed
elongation steps were not found within the ped cluster.
several unprecedented features among type I PKSs. Neverthe-
Unclustered genes of a bacterial PKS pathway are very rare: to
less, its unusual architecture clearly mirrors the structure of the
the best of our knowledge, the ansamitocin system is the only
pederin (1) starter moiety. The deduced product of pedI
other published example of a disconnected PKS.[19] To isolate the
resembles a type I PKS consisting of four modules. By phyloge-
remaining pederin genes from the bacterial consortium of
netic analysis of a PCR product covering a KS domain of this
P. fuscipes, we used a phylogeny-based PCR strategy that
gene, we had previously predicted that the PKS lacks AT
specifically targets trans-AT PKS systems.[18] This approach led
domains. In accordance with this, no homology to such domains
to the isolation of additional PKS gene fragments from
was found within PedI. The first module lacks a KS domain and
metagenomic DNA of beetle bacteria. In this paper we present
should therefore catalyze chain initiation. However, instead of a
evidence that one of these fragments belongs to the missing PKS
conventional AT, the module harbors an unusual N-terminal
catalyzing the first three rounds of elongation.
domain of about 240 aa similar to proteins of the GCN5-relatedN-acetyltransferase (GNAT) family[20] and sequence patternsdiagnostic
[RQ]XXGX[GA] motif involved in binding of the acetyl-CoAsubstrate[20] was identified at aa 135 ± 140 (QQYNLG). GNATs
Screening of a bacterial cosmid library for additional trans-AT
catalyze the transfer of an acetyl group from acetyl-CoA to
various heteroatoms,[20] which suggests that this domain primes
We have previously reported the PCR amplification from
the loading module with acetate. GNAT domains as part of PKSs
metagenomic bacterial DNA of three PKS gene fragments
have not been reported before.[9, 21] All published AT-less PKS
(PS1 ± PS3) that do not belong to the already sequenced ped
systems are loaded with a starter other than acetate. In these
cluster.[18] PCR products covering the KS domain of trans-AT PKSs
cases, the absence of a GNAT domain, which usually accepts only
¹ 2004 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Figure 1. Map of the genomic region belonging to the PCR product PS1 and proposed biosynthesis of pederin (1). Dotted boxes and arrows indicate degraded ORFs. Pseudogenes in black are involved in horizontal gene transfer. Ped genes are marked in gray. GNAT, GCN5-related N-acetyltransferase domain; ACP, acyl carrier proteindomain; KS, ketosynthase domain; KR, ketoreductase domain; MT, methyltransferase domain; CR, crotonase domain; OXY, oxygenase; T, transposase; INT, phageintegrase. Numbered genes are highly similar to genes from Pseudomonas aeruginosa. The numbers refer to the PA number of the closest homologue. PedA ± PedH areencoded on a different part of the symbiont genome.[7]
acetyl-CoA as substrate, is therefore not surprising. The
that were proposed to catalyze the addition of acetyl- or
sequencing of additional AT-less PKSs will reveal if the PedI
malonyl-CoA to a keto group, followed by decarboxylation,
GNAT domain is an exception or a common feature to acetate-
dehydration, and/or reduction steps. We had initially expected a
similar enzyme in the biosynthesis of pederin (1), but such a
In the second module, signature motifs of a KS, a ketoreduc-
homologue is not among the genes on pPS9D2 and could
tase (KR), a methyltransferase (MT), and an ACP domain were
neither be detected in the symbiont DNA by hybridization with a
found, indicating that the corresponding polyketide extension
probe derived from the HMG-CoA synthase homologue mupH
unit bears a methyl group in an a-position to a hydroxy function.
nor by PCR with degenerate primers based on conserved motifs
The KR domain contains a characteristic Asp residue at
(data not shown). Instead, two unprecedented domains were
position 1270, which has recently been shown to be conserved
found in the third module of PedI. In addition to a KS and an ACP
in domains generating alcohols with D stereochemistry[22] (B-type
domain, two domains were identified that show homology to
alcohols, as defined in ref. [23]). This precisely corresponds to the
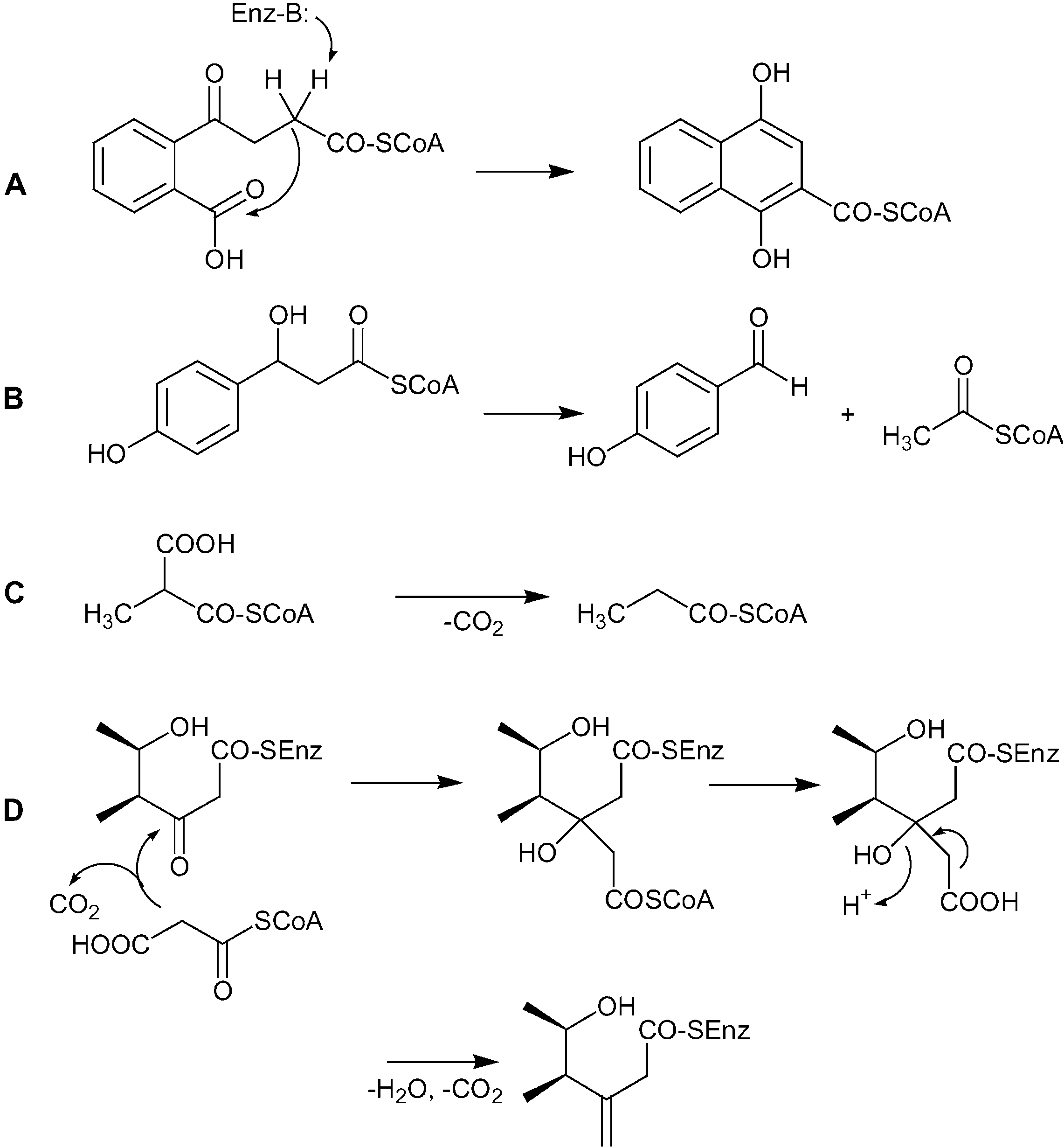
enzymes of the crotonase (CR) superfamily. These enzymes
first extension unit of the pederin molecule, which bears a B-type
catalyze a wide range of diverse reactions all involving a
hydroxyl function adjacent to a methyl group. Cardani et al.
stabilized CoA thioester enolate intermediate.[28] The CR domain
attempted to elucidate the biosynthesis of 1 by feeding
adjacent to the KS exhibits relatively low homology to known
radioactively labeled precursors to P. fuscipes.[24] They reported
enzymes (42 % similarity to the next homologue). A PSI-BLAST
the presence of 40 % activity of administered labeled propionate
search revealed that it is related to naphthoate synthases.[29] The
in the N-acyl moiety constituting the western half of pederin (1).
second CR domain is highly similar to discrete enzymes found in
This seemingly suggests that the first extension unit is derived
trans-AT systems, such as PksI (66 %) from Bacillus subtilis[15] and
from methylmalonyl-CoA instead of malonyl-CoA and SAM-
MupK from Pseudomonas fluorescens.[13] These enzymes are
dependent methylation by an MT domain. However, propionate
annotated as putative enoyl-CoA hydratases, but their exact
is catabolized in many bacteria[25] and insects[26] to acetate by
functions are presently unknown. If the colinearity rules apply,
several different routes and can then enter polyketide biosyn-
this module could catalyze the attachment of the exomethylene
thesis. Since the radioactive label was located on the propionate
group, possibly by action of the two unusual CR domains.
C2 position, it would be retained after chain shortening and be
Indeed, members of the CR superfamily perform reactions very
incorporated into malonyl-CoA-derived moieties. The fact that
similar to those proposed for exomethylene group formation
all known methyl groups generated by AT-less PKSs are attached
(Scheme 1). Among the enzymes acting on CÀC bonds, naph-
by MT domains[12, 13, 15] is consistent with this hypothesis.
thoate synthase from menaquinone metabolism catalyzes a
The second extension unit bears a rare exomethylene group at
Claisen condensation,[29] and hydroxycinnamate lyase from
its C1 position. Carbon atoms that branch off such positions are
Ps. fluorescens is responsible for a retro-aldol reaction.[30] Decar-
known from a number of other complex polyketides, for
boxylation and dehydration steps are known from the E. coli
example, myxovirescin ( TA),[27] mupirocin,[13] and leinamycin.[12]
methylmalonyl-CoA decarboxylase and crotonase,[28] respective-
The PKS clusters of these compounds encode 3-hydroxy-3-
ly. Moreover, homologues of the second CR domain are located
methylglutaryl coenzyme A (HMG-CoA) synthase homologues
directly adjacent to HMG-CoA synthase-like genes in the mup
¹ 2004 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Table 1. Genes and pseudogenes identified in the PS1 region.
GNAT, GCN5-related histone acetyltransferase family
branched-chain amino acid transporter permease protein
branched-chain amino acid transporter permease protein
branched-chain amino acid transporter ATP-binding protein
branched-chain amino acid transporter ATP-binding protein
[a] Entries in parentheses belong to pseudogenes.
and pksX clusters, suggesting that these genes participate in
N-terminal KS domain of PedF lacks the conserved His residue
methyl(ene) group formation. The first CR domain without close
necessary for the decarboxylation of the extender unit.[31] This
homologues might substitute the HMG-CoA synthase, catalyzing
module should therefore be unable to perform a polyketide
the aldol addition of acetyl- or malonyl-CoA (Scheme 1). The CR
elongation, which in fact necessitates an additional module on
related to PksI and MupK could then be involved in the further
PedI. The same feature is known from the mup system, in which a
processing of the intermediate and perform the final decarbox-
seemingly superfluous module lacking a decarboxylation motif
ylation and/or dehydration steps. If the two CR domains are
is located at the N-terminus of MmpA.[13] Thomas and co-workers
indeed sufficient for exomethylene group formation, this
proposed that such modules could have a pseudo-loading
module could represent a valuable tool in engineered biosyn-
function and be involved in the proper channeling of polyketide
thesis that might dramatically increase the possible structural
chains between two PKS proteins.[13] Alignment of the last PedI
range of recombinant polyketides. However, at present it cannot
module with other PKSs (data not shown) revealed that the KS
be excluded that PedK or additional enzymes encoded else-
and KR domains are separated by an unusually large region of
where on the symbiont genome participate in this reaction.
approximately 250 aa without convincing homology to known
Module 4 represents the last module of this PKS and contains
sequences or patterns. Further functional studies are needed to
a KS, a KR, and an ACP domain. The presence of this module was
determine whether it represents a novel domain, perhaps
unexpected, since the corresponding polyketide extension step
catalyzing the stereospecific intramolecular acetalization that
was initially assigned to the first module of PedF. However, the
likely takes place at this point in the biosynthesis of pederin (1),
¹ 2004 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
needs additional genes to be functional. These featuresstrongly suggest that PedIJK and the previously identi-fied ped genes complement each other and jointlyencode pederin (1) biosynthesis. The current uncultur-ability of the Paederus spp. symbiont prohibits standardexperiments to verify the function of ped genes, such asknockout. Present studies in our laboratory thereforeaim at expressing the ped genes in a heterologous host.
Although the ped system is to the best of our
knowledge only the second example of PKS genesdistributed among two genomic regions,[19] the numer-ous IS elements located on both ends of the two pedregions readily explain the disjointed structure. Theirpresence indicates that the entire PKS cluster wasoriginally acquired as a genomic island by horizontalgene transfer. By a second IS-mediated rearrangementprocess, the pedIJK region could then have transposedto a different genome position. Ultimately, the regionwould have been stabilized by decay of all adjacentmobility genes. In free-living bacteria, frequent horizon-tal gene transfer selects for the clustering of genes fromsecondary metabolism.[33] In contrast, gene exchangewould be a rare event in the Paederus spp. symbiont,which has lived in a shielded environment for millions ofyears. These conditions could have resulted in theobserved gene scattering.
Scheme 1. Reactions catalyzed by members of the crotonase family and hypothetical
The pederin system represents an ideal model to
pathway leading to the formation of the pederin exomethylene group. A) napthoate
understand the factors governing natural product
synthase (MenB); B) 4-hydroxycinnamoyl-CoA hydratase/lyase (HCHL) from Ps. fluorescensAN103 (last step of a two-step reaction); C) methylmalonyl-CoA decarboxylase (MMCD)
symbioses and how sustainable sources of drug candi-
from E. coli; D) proposed pathway catalyzed by the PedI CRs. Acetyl-CoA could be used
dates from invertebrates can be created. In this work,
instead of malonyl-CoA in this route.
the availability of all gene candidates necessary forpederin (1) biosynthesis now sets the stage for theproduction of pederin-type antitumor compounds in a
or whether it is only an unusually long spacer region between
culturable bacterium. In addition, the small repertoire of known
the KS and the KR. The KR domain present in this module has no
PKS domains has been expanded by at least three new members.
structural counterpart in pederin (1). It could be an inactive
Their further study could significantly improve the versatility of
evolutionary remnant, similar to many other known PKS domains
engineered biosynthesis and our understanding of polyketide
with no apparent function.[9] Alternatively, it could aid in the
acetalization by polarizing the carbonyl bond.
PedJ, encoded by a gene downstream of pedI, is highly similar
to several oxidoreductases of the bacterial luciferase type and
contains the characteristic Pfam pattern (PF00296). The twoclosest database matches are MupA with unknown function[13]
Isolation of bacteria from Paederus fuscipes: One hundred
and MtaG from the myxothiazol pathway.[32] The latter enzyme
P. fuscipes beetles collected at Aydın, Turkey were ground in liquid
was proposed by M¸ller and coworkers to catalyze the a-
nitrogen and resuspended in LB medium (5 mL). The mixture was
hydroxylation of an amino acid residue. Mupirocin contains no
kept for 10 min on ice to let beetle residues settle. The supernatant
amino acid but features an a-hydroxylated polyketide extender
was centrifuged at 100 g for 10 min to sediment eukaryotic cells. The
position. PedJ could therefore be responsible for the a-
supernatant was then passed through a 40-mm nylon filter (Milli-
hydroxylation of either the glycine residue or the third extension
pore), and the bacteria were pelleted by centrifugation at 5000 g for10 min. The purity of the bacterial fraction was checked by DAPI
unit. Adjacent to pedJ, the ORF pedK encoding a putative protein
staining and microscopic inspection.
of 318 aa was identified. However, homology and patternanalysis provided no insight into the possible function of this
Preparation and screening of a P. fuscipes symbiont genomiclibrary: Genomic DNA from P. fuscipes bacteria was prepared by
standard SDS lysis and used for the construction of a cosmid library
In contrast to all other known PKS gene clusters, no domain or
in the pWEB vector (Epicentre) according to the manufacturer's
gene was identified on the cosmid that would be involved in
instructions. Two thousand clones were deposited in 96-well plate
release of the assembled chain from the PKS or in the transfer of
format and transferred to Hybond N membranes. Membranes were
acyl-CoA units to the ACP domains. Therefore, this PKS evidently
screened by using a fluorescein-labeled probe prepared from a 1.5-
¹ 2004 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
kb fragment of the ped cluster and the ECL Direct Labeling and
[4] P. Proksch, R. A. Edrada, R. Ebel, Appl. Microbiol. Biotechnol. 2002, 59, 125.
Detection kit (Amersham Biosciences) according to the manufac-
[5] R. Narquizian, P. J. Kocienski in The Role of Natural Products in Drug
turer's protocol, except that Church buffer was used during
Discovery, Vol. 32 (Eds.: R. Mulzer, R. Bohlmann), Springer, Heidelberg,
prehybridization and hybridization. The probes were prepared from
[6] R. L. L. Kellner, K. Dettner, Oecologia 1996, 107, 293.
PCR fragments corresponding to the KS domain as described
[7] J. Piel, Proc. Natl. Acad. Sci. USA 2002, 99, 14 002.
elsewhere.[7, 18] Hybridization was performed at 60 8C, followed by
[8] R. L. L. Kellner, Insect Biochem. Mol. Biol. 2002, 32, 389.
low-stringency washes for 2 Â 7 min at 25 8C and 2 Â 15 min at 60 8C.
[9] B. J. Rawlings, Nat. Prod. Rep. 2001, 18, 231.
Sequence analysis of positive cosmids: Inserts of positive cosmids
[10] E. Rodriguez, R. McDaniel, Curr. Opin. Microbiol. 2001, 4, 526.
were end-sequenced to gain information about the location of PKS
[11] C. D. Reeves, Crit. Rev. Biotechnol. 2003, 23, 95.
genes. The cosmid pPS9D2 with genes unrelated to PKSs at its
[12] Y. Q. Cheng, G. L. Tang, B. Shen, Proc. Natl. Acad. Sci. USA 2003, 100, 3149. [13] A. K. El-Sayed, J. Hothersall, S. M. Cooper, E. Stephens, T. J. Simpson, C. M.
termini was chosen for complete sequencing. After shearing of the
DNA by using a Standard Nebulizer (Octurno), the fragment ends
[14] S. Mochizuki, K. Hiratsu, M. Suwa, T. Ishii, F. Sugino, K. Yamada, H. Kinashi,
were repaired with T4 DNA polymerase and Klenow fragment.
Fragments of 1 ± 1.5 kb were isolated by agarose electrophoresis,
[15] A. M. Albertini, T. Caramori, F. Scoffone, C. Scotti, A. Galizzi, Microbiology
cloned into the pUC18 vector, and end-sequenced by using the
BigDye Terminator Ready Mix (Applied Biosystems) and an ABI 3700
[16] Y. Paitan, G. Alon, E. Orr, E. Z. Ron, E. Rosenberg, J. Mol. Biol. 1999, 286, 465.
sequencer (Applied Biosystems). Sequence data were assembled by
[17] G. Z. Huang, L. H. Zhang, R. G. Birch, Microbiology 2001, 147, 631.
using GAP4 software[34] and analyzed by using the BLASTX, PSI-
[18] J. Piel, D. Hui, N. Fusetani, S. Matsunaga, Environ. Microbiol., in press.
BLAST, FramePlot, and InterProScan algorithms.
[19] T. W. Yu, L. Q. Bai, D. Clade, D. Hoffmann, S. Toelzer, K. Q. Trinh, J. Xu, S. J.
Moss, E. Leistner, H. G. Floss, Proc. Natl. Acad. Sci. USA 2002, 99, 7968.
The nucleotide sequence has been deposited at GenBank under
[20] A. F. Neuwald, D. Landsman, Trends Biochem. Sci. 1997, 22, 154.
[21] B. S. Moore, C. Hertweck, Nat. Prod. Rep. 2002, 19, 70. [22] R. Reid, M. Piagentini, E. Rodriguez, G. Ashley, N. Viswanathan, J. Carney,
D. V. Santi, C. R. Hutchinson, R. McDaniel, Biochemistry 2003, 42, 72.
[23] P. Caffrey, ChemBioChem 2003, 4, 654.
[24] C. Cardani, C. Fuganti, D. Ghiringh, P. Grassell, M. Pavan, M. Valcuron,
We thank H¸seyin BasÀpınar and Ekin SÀavk for helping with the
[25] S. Textor, V. F. Wendisch, A. DeGraaf, U. M¸ller, M. I. Linder, D. Linder, W.
collections of Turkish beetle specimens, Elke Meier, Kathleen Seitz,
Buckel, Arch. Microbiol. 1997, 168, 428.
[26] P. P. Halarnkar, J. D. Chambers, G. J. Blomquist, Comp. Biochem. Physiol.
Ivonne Hˆfer, and Domenica Schnabelrauch for technical assis-
Part B: Biochem. Mol. Biol. 1986, 84, 469.
tance, and Konrad Dettner for valuable discussions. We are
[27] Y. Paitan, E. Orr, E. Z. Ron, E. Rosenberg, Microbiology 1999, 145, 3059.
indebted to Chris M. Thomas for providing the mupirocin producer
[28] H. M. Holden, M. M. Benning, T. Haller, J. A. Gerlt, Acc. Chem. Res. 2001, 34,
Ps. fluorescens NCIMB 10586, and to Wilhelm Boland for support.
[29] V. Sharma, K. Suvarna, R. Meganathan, M. E. S. Hudspeth, J. Bacteriol.
This work was supported by Research Grants from the Deutsche
Forschungsgemeinschaft (PI 430/1-1 and PI 430/2-1) and the Max
[30] M. J. Gasson, Y. Kitamura, W. R. McLauchlan, A. Narbad, A. J. Parr, E.
Lindsay, H. Parsons, J. Payne, M. J. C. Rhodes, N. J. Walton, J. Biol. Chem. 1998, 273, 4163.
[31] A. Witkowski, A. K. Joshi, S. Smith, Biochemistry 2002, 41, 10 877.
Keywords: biosynthesis ¥ natural products ¥ polyketides ¥
[32] B. Silakowski, H. U. Schairer, H. Ehret, B. Kunze, S. Weinig, G. Nordsiek, P.
symbiosis ¥ synthases ¥ uncultivated bacteria
Brandt, H. Blocker, G. Hˆfle, S. Beyer, R. M¸ller, J. Biol. Chem. 1999, 274,37 391.
[33] J. G. Lawrence, J. R. Roth, Genetics 1996, 143, 1843. [34] J. K. Bonfield, K. F. Smith, R. Staden, Nucl. Acids Res. 1995, 23, 4992.
[1] J. Kobayashi, M. Ishibashi, Chem. Rev. 1993, 93, 1753. [2] J. Faulkner, M. D. Unson, C. A. Bewley, Pure Appl. Chem. 1994, 66, 1983.
[3] M. G. Haygood, E. W. Schmidt, S. K. Davidson, J. D. Faulkner, J. Mol.
¹ 2004 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Identificación. en la UC: 7548872-2880970 Casa: 4537153 Consulta: 2078372-2060668 Celular (09): 2241711 Estudios Universitarios de Pregrado. : Medicina, Universidad de Concepción y Universidad de Chile, Chile Formación de Postitulo o de Postgrado. : Becado de Retorno en Psiquiatría. Hospital del Salvador. Servicio de Salud Metropolitano Oriente. Santiago, Abril de 1982 - Nov
BAKGRUNN Nasjonalt Kompetansesenter for Søvnsykdommer (SOVno) ble etablert i september 2004, og er lokalisert til Psykiatrisk Klinikk på Haukeland Universitetssykehus. Det er Helsedepartementet som fatter vedtak om etablering av Nasjonale kompetansesentra. SOVno skal sikre nasjonal kompetanseoppbygging og kompetansespredning innenfor søvn. Viktige oppgaver er (se for øvrig http://www.dep.no/

 Unprecedented Diversity of Catalytic Domains inthe First Four Modules of the Putative PederinPolyketide SynthaseJˆrn Piel,*[a] Gaiping Wen,[b] Matthias Platzer,[b] and Dequan Hui[a]
Polyketides of the pederin group are highly potent antitumor
region of the symbiont genome. It contains at least three novel
compounds found in terrestrial beetles and marine sponges.
Unprecedented Diversity of Catalytic Domains inthe First Four Modules of the Putative PederinPolyketide SynthaseJˆrn Piel,*[a] Gaiping Wen,[b] Matthias Platzer,[b] and Dequan Hui[a]
Polyketides of the pederin group are highly potent antitumor
region of the symbiont genome. It contains at least three novel
compounds found in terrestrial beetles and marine sponges.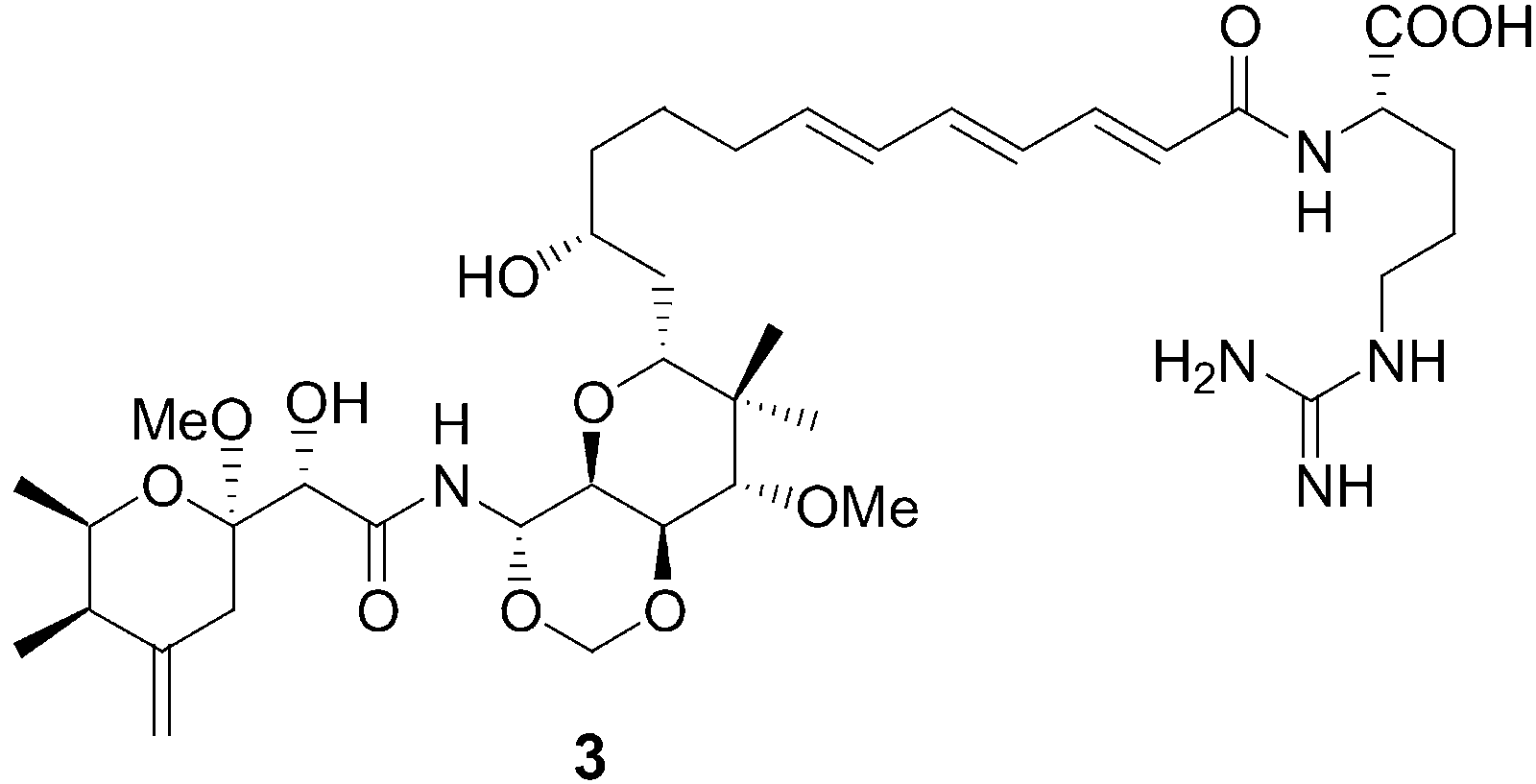 can be distinguished from those of cis-AT systems by phyloge-netic analysis.[18] Such an analysis placed the amplified productdesignated as PS3 close to the cis-AT, and PS1 and PS2 into thetrans-AT group. Since the missing part of the ped PKS shouldbelong to the latter group, we intended to isolate the gene(s)belonging to these PCR products. We therefore screened2000 clones of a cosmid library prepared from DNA of P. fuscipesbacteria with probes derived from PS1 and PS2. Unexpectedly,restriction mapping of the positive clones revealed that the twoPCR products did not originate from the same genomic region
an acyl carrier protein (ACP) domain, and attaches one polyke-
and apparently belonged to different secondary metabolites.
can be distinguished from those of cis-AT systems by phyloge-netic analysis.[18] Such an analysis placed the amplified productdesignated as PS3 close to the cis-AT, and PS1 and PS2 into thetrans-AT group. Since the missing part of the ped PKS shouldbelong to the latter group, we intended to isolate the gene(s)belonging to these PCR products. We therefore screened2000 clones of a cosmid library prepared from DNA of P. fuscipesbacteria with probes derived from PS1 and PS2. Unexpectedly,restriction mapping of the positive clones revealed that the twoPCR products did not originate from the same genomic region
an acyl carrier protein (ACP) domain, and attaches one polyke-
and apparently belonged to different secondary metabolites.
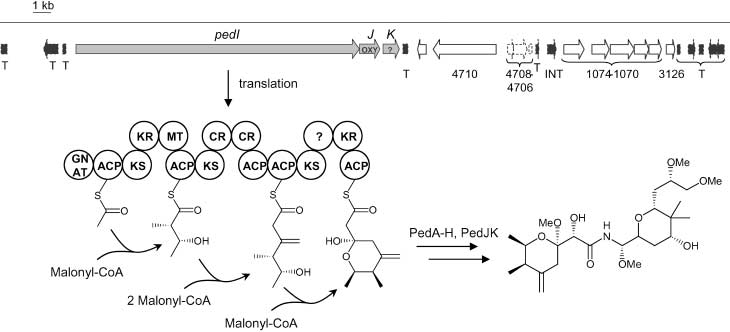 Figure 1. Map of the genomic region belonging to the PCR product PS1 and proposed biosynthesis of pederin (1). Dotted boxes and arrows indicate degraded ORFs.
Figure 1. Map of the genomic region belonging to the PCR product PS1 and proposed biosynthesis of pederin (1). Dotted boxes and arrows indicate degraded ORFs.
 needs additional genes to be functional. These featuresstrongly suggest that PedIJK and the previously identi-fied ped genes complement each other and jointlyencode pederin (1) biosynthesis. The current uncultur-ability of the Paederus spp. symbiont prohibits standardexperiments to verify the function of ped genes, such asknockout. Present studies in our laboratory thereforeaim at expressing the ped genes in a heterologous host.
needs additional genes to be functional. These featuresstrongly suggest that PedIJK and the previously identi-fied ped genes complement each other and jointlyencode pederin (1) biosynthesis. The current uncultur-ability of the Paederus spp. symbiont prohibits standardexperiments to verify the function of ped genes, such asknockout. Present studies in our laboratory thereforeaim at expressing the ped genes in a heterologous host.