Thank you, it's cool. Used for the first time because only then was the right drug. Very revealing detail: ordered Express shipping to get the order within 3 hours because didn't know how to get the order faster https://africarx.co.za/ The drug is authentic exactly. Consistent with the stated prices the Staff is knowledgeable.
Unescientificwritingseminar.wikispaces.com
Prefrontal Mechanisms in Extinction of Conditioned Fear Gregory J. Quirk, René Garcia, and Francisco González-Lima Interest in the medial prefrontal cortex (mPFC) as a source of behavioral inhibition has increased with the mounting evidence for afunctional role of the mPFC in extinction of conditioned fear. In fear extinction, a tone-conditioned stimulus (CS) previously pairedwith a footshock is presented repeatedly in the absence of footshock, causing fear responses to diminish. Here, we review convergingevidence from different laboratories implicating the mPFC in memory circuits for fear extinction: (1) lesions of mPFC impair recallof extinction under various conditions, (2) extinction potentiates mPFC physiological responses to the CS, (3) mPFC potentiation iscorrelated with extinction behavior, and (4) stimulation of mPFC strengthens extinction memory. These findings support Pavlov’soriginal notion that extinction is new learning, rather than erasure of conditioning. In people suffering from posttraumatic stressdisorder (PTSD), homologous areas of ventral mPFC show morphological and functional abnormalities, suggesting that extinctioncircuits are compromised in PTSD. Strategies for augmenting prefrontal function for clinical benefit are discussed.Key Words: Amygdala, infralimbic, long-term potentiation, prelim-
in rodents supporting this hypothesis. We also suggest ways in
which prefrontal mechanisms of extinction may be augmentedso as to enhance extinction, with potential clinical applications.
Thestudyoffearandanxietyinexperimentalanimalshas LesionStudies
advanced rapidly with the use of Pavlovian fear condition-ing, in which a tone-conditioned stimulus (CS) is associ-
The idea that extinction circuits involved the prefrontal cortex
ated with a footshock unconditioned stimulus (US). Conditioned
originated with early primate studies of appetitive conditioning,
fear reactions to the tone extinguish in the absence of the shock.
in which lesions of the ventral mPFC (vmPFC) and orbitofrontal
The resurgence of interest in extinction is due in large part to its
cortex resulted in increased responding during extinction
potential applicability to the treatment of anxiety disorders, such
for a complete history of prefrontal cortex in extinc-
as posttraumatic stress disorder (PTSD), in which extinction is
thought to be compromised. A thorough understanding of the
neural circuits of extinction of fear could yield new treatments for
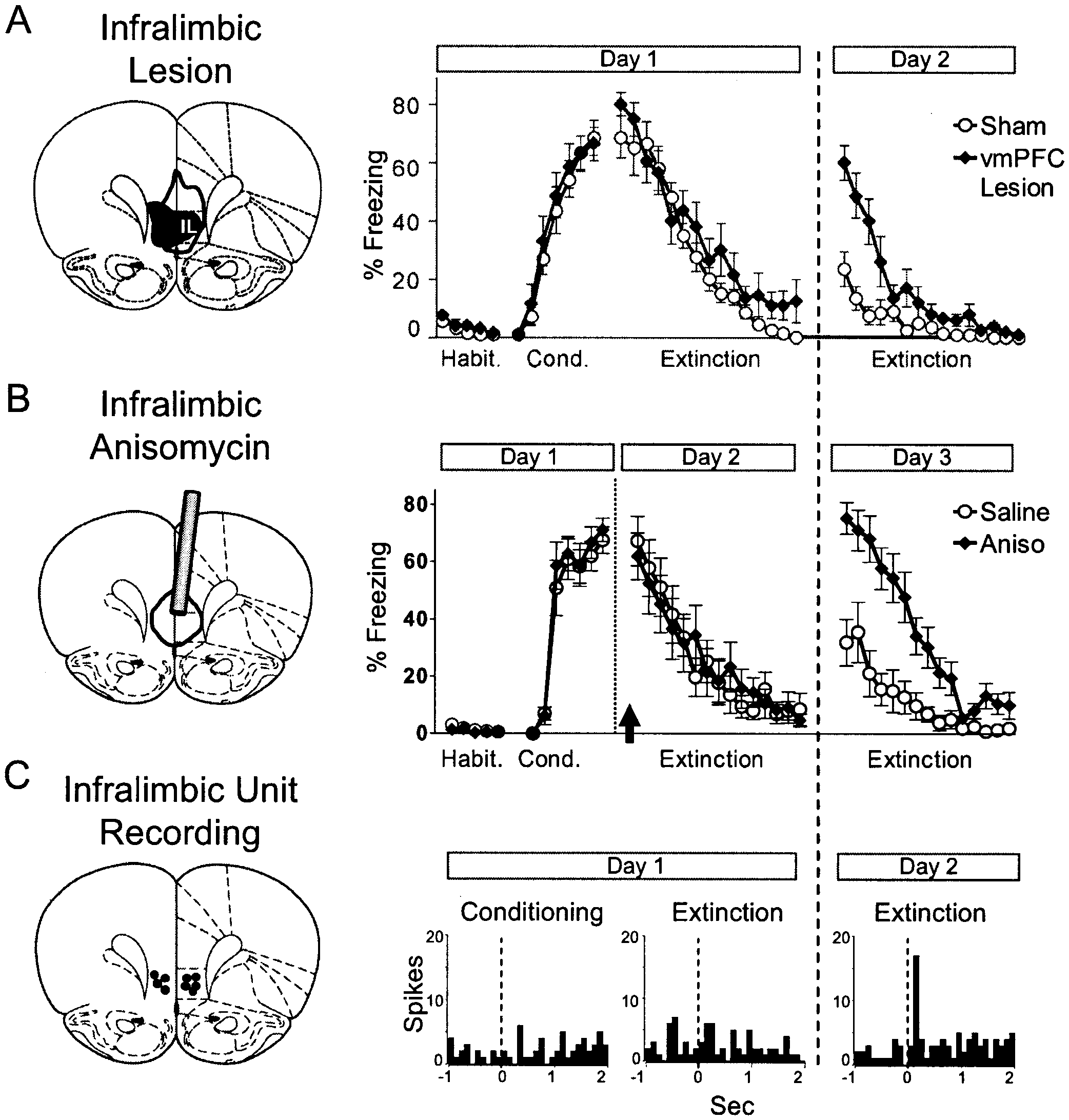
observed that rats with vmPFC lesions could acquire fear nor-
augmenting exposure-based therapies that are used to treat
mally but had difficulty extinguishing across several days of
In his classic investigation of appetitive conditioning in dogs,
that rats with vmPFC lesions that were centered on the infralim-
Pavlov observed that extinguished responses spontaneously
bic cortex (IL) could extinguish normally within a session but
recovered with the passage of time This suggested
had difficulty recalling extinction 24 hours later, suggesting that
that extinction did not erase the memory for conditioning but
IL is not required for fear inhibition under all circumstances but
represented new learning. More recent behavioral studies have
is important for recalling extinction after a long delay. Other
confirmed and extended this finding for conditioned fear
studies have confirmed that vmPFC lesions impair recall of
extinction does not erase the conditioning memory, it must form
a new memory that inhibits the conditioned response. This
suggests that some structure or structures are activated byextinction, so as to excite inhibitory circuits that are responsible
Recording Studies
for reducing the expression of fear Despite early
Lesion studies presuppose that regional contributions to brain
theoretical formulations of extinction-related inhibition
function may be inferred from a damaged brain. A more direct
the search for inhibitory circuits largely
approach is to record from neuronal activity in awake animals
undergoing extinction training. Do mPFC neurons signal extinction?
However, studies that build on recent advances in the
Paralleling mPFC lesion findings, single neurons in IL did not signal
acquisition of conditioned fear point to the medial prefrontal
the tone CS during acquisition or extinction training
cortex (mPFC) as an important part of the neural circuit for fear
The next day, however, when rats were recalling
extinction. In this review, we describe converging evidence from
extinction, IL units showed potentiation of short-latency tone re-
lesion, recording, metabolic, stimulation, and microinfusion studies
sponses The larger the tone response, the lower thespontaneous recovery of freezing, consistent with IL-mediatedinhibition of fear after extinction. No such potentiation was ob-
From the Department of Physiology (GJQ), Ponce School of Medicine,
served in adjacent prelimbic cortex. Thus, extinction potentiated
Ponce, Puerto Rico; Neurobiologie et Psychopathologie (RG), Universite
auditory inputs to IL neurons, providing direct support for the
de Nice-Sophia Antipolis, Nice, France; and the Institute for Neuro-
Pavlov-Konorski hypothesis that extinction potentiates neuronal
science and Department of Psychology (FGL), University of Texas at
activity in structures that are involved in inhibition of the condi-
Address reprint requests to Gregory J. Quirk, Ph.D., Department of Physiol-
ogy, Ponce School of Medicine, P.O. Box 7004, Ponce, Puerto Rico 00732;
What inputs to mPFC might become potentiated as a result of
Received August 1, 2005; revised October 16, 2005; accepted March 3, 2006.
whether repeated presentations of a tone CS in the absence of
doi:10.1016/j.biopsych.2006.03.010 2006 Society of Biological Psychiatry
behavioral networks. Metabolic responses to a test tone were
compared in groups of mice that received fear conditioning, a
pseudorandom treatment (unpaired tones and shocks), or con-
ditioning followed by extinction. Consistent with single-unit andevoked-potential recording, the largest increase in metabolic activityafter extinction occurred in the mPFC. The infralimbic (but not theprelimbic) area showed significantly more metabolic activity thancontrols. In addition to IL, significant metabolic increases were
Conditioning Extinction
observed in dorsal, medial, and lateral frontal cortex, which areareas not yet studied with the unit-recording technique. Hence,
Figure 1. Schematic relating conditioned behavior to memory for condi-
multiple prefrontal regions may play a role in extinction memory.
tioning and extinction. As first suggested by Pavlov, extinction training does
There also were changes in the interaction between the prefron-
not eliminate memory for conditioning but generates a new memory that
tal cortex and other regions, particularly in auditory and limbic
competes with conditioning for control of behavior. For conditioned fear,this schema suggests that there are structures in the brain that increase their
networks. In support of an inhibitory role, FDG labeling in
neuronal activity with extinction, so as to drive down fear via inhibition of
dmPFC, in IL cortex, and in dorsal and lateral frontal cortex was
correlated significantly with extinction behavior Finally, there was a strong negative correlation between
the US induces long-term potentiation (LTP) in the mPFC. The
prefrontal areas and regions thought to be involved in expression
mPFC receives glutamatergic inputs from the hippocampus
of conditioned fear, such as the ventral tegmental area, MD
thalamus, and the entire auditory system (brainstem, thalamic,
High-frequency stimulation of each of these input areas results in
These mapping data suggest that extinction training engages
a network of interactive brain regions, which may serve two
functions: to inhibit the conditioned response after extinction
pathways have confirmed development of LTP-like changes in
and to preserve some of the original CS-US associative effects
the mPFC with extinction training. MD-evoked responses inmPFC show little change during extinction training but areincreased 1–7 days after extinction Similarly, extinction-related LTP takes place in thehippocampal–mPFC pathway after extinction training Interestingly, failure to recall extinction wasassociated with inhibition of MD-evoked potentials, and depressingthe MD-mPFC pathway with low-frequency stimulation caused fullrecovery of conditioned fear after extinction Thus, extinction training results in LTP of thalamicinputs to mPFC even days after extinction, paralleling the single-unitrecording studies and indicating a role ofmPFC in long-term retention of extinction memory. Thus, inputs tothe mPFC from the thalamus, hippocampus, or the BLA maybecome potentiated after extinction.
Microinfusion data have strongly implicated the BLA in acqui-
sition of extinction however, lesions of the basal nucleus have no effect on short- orlong-term memory for extinction This highlights the potentialimportance of the lateral amygdala in the acquisition of extinc-tion. Another potentially important input to mPFC is the auditoryassociation cortex in light of reports thatauditory-cortex lesions impair extinction of auditory fear condi-tioning Local inactivationof various inputs to mPFC is needed to determine which onesmay impair extinction learning and memory. Figure 2. Converging lines of evidence showing that the infralimbic pre- Metabolic Mapping
frontal cortex (IL) is functionally involved in recall of extinction. (A) Lesions of IL do not prevent extinction but interfere with recall of extinction the follow-
In addition to single-unit and evoked potential recording,
ing day (modified from Quirk et al 2000). (B) Unit recording shows that IL
extinction of auditory conditioning also has been investigated
neurons respond to the tone only during recall of extinction, suggesting
with metabolic-mapping techniques that assess the uptake of
that IL tone responses are responsible for low fear after extinction (modified
fluorodeoxyglucose (FDG), a radiolabeled glucose analog
from Milad and Quirk, 2002). (C) Infusing the protein synthesis inhibitor
Brain activity can be mapped with FDG because
anisomycin (Aniso) into the IL just before extinction (arrow) has no effect onextinction learning but blocked recall of extinction the following day (mod-
brain cells use glucose and its analogs for energy metabolism
ified from Santini et al 2004). These and other data suggest that extinction-
An important advantage of metabolic mapping
induced potentiation of prefrontal neuronal activity is necessary for sup-
over electrophysiological recording methods is that the entire
pression of fear after extinction. vmPFC, ventral medial prefrontal cortex;
brain can be examined at once, permitting visualization of
Habit., habituation; Cond., conditioning. Table 1. Converging Lines of Evidence from Recent Rodent Studies Showing that Extinction can be Facilitated by Activation of Medial Prefrontal Cortex (mPFC)
Electrical stimulation paired with CSa,e
Field potentials evoked by thalamic stimulationb
Field potentials evoked by thalamic stimulationd
Long-term potentiation of thalamic inputsd
Metabolic enhancement with methylene bluef
from acquisition. Thus, there is remarkable convergence be-
memory, although transcription inhibitors and transgenic ap-
tween the three different techniques (single-unit, evoked poten-
proaches will be needed to determine whether gene expression
tial, metabolic mapping) in two species (rat and mouse), show-
ing that extinction potentiates vmPFC responses to the tone CS These results clearly support Pavlov’s cortical inhibi-
Expression of Extinction
tion hypothesis and contradict the simpler notions of extinction
Once potentiated, how does mPFC inhibit fear after extinc-
as unlearning or reversal of acquisition.
tion? The infralimbic subregion of mPFC has extensive projec-tions to the amygdala, as well as the amygdala’s targets in the
Molecular Studies
Formation of long-term memory has been linked to a molec-
If these projections are inhibitory, the IL could
ular cascade involving N-methyl-D-aspartate (NMDA)-mediated
override amygdala-generated fear responses. The physiological
calcium entry, activation of protein kinases, gene expression,
effect of many of these projections is not known, but anatomical
and protein synthesis Involvement of this cascade
support exists for IL-mediated inhibition of the amygdala. IL
in extinction would provide support for the idea that extinction
projects robustly to the region between the central and basolat-
constitutes new learning. It has been known for some time that
eral nuclei, containing intercalated (ITC) cells
formation of long-term memory for extinction. Protein kinases
responsible for feed-forward inhibition of central nucleus output
and protein synthesis in the amygdala also have been implicated
neurons In support of this model, electrical
stimulation of the IL area decreased the excitability of brainstem-
Recent evidence suggests that a similar molecular cascade oper-
projecting neurons of the amygdala central nucleus
ates in the mPFC during extinction. Antagonists of NMDA receptors
and decreased the expression of conditioned fear
According to this model extinction-induced
potentiation of tone responses in IL neurons would cause
prevent the formation of long-term (but not short-term) extinction
feed-forward inhibition of the central nucleus, thereby prevent-
when microinfused into the mPFC. In each case, delaying the
ing fear signals in BLA from exiting the amygdala. Consistent
infusion 2 or 4 hours after extinction eliminated the effect, consistent
with this, it recently was shown that chemical stimulation of IL
with a time-limited role of molecular processes in consolidation ofextinction. Western blot analysis of prefrontal tissue shows that
A. Before Extinction B. After Extinction
infusion of MAPk inhibitor PD098059 into the mPFC immediatelyafter extinction decreased levels of phosphorylated ERK2 without
affecting total ERKs Future experiments will
determine whether inhibition of extracellular signal-regulated ki-
nase-2 (ERK2) phosphorylation is related to LTP in the MD-mPFC or
hippocampal–mPFC pathways or to other inputs to the mPFC (for
Does activation of the ERK-MAPk system in the mPFC trigger
gene expression necessary for extinction memory? Although little
Figure 3. Schema for mPFC inhibition of fear via the amygdala. (A) Before
is known about extinction-induced gene expression, it recently
extinction, the tone CS activates the basolateral amygdala (BLA), which
was shown that extinction training stimulates the immediate
activates the central nucleus (Ce) output neurons, triggering fear responses.
(B) After extinction, prefrontal (PFC) responses to the tone are potentiated,
Controls indicated that this up-regulation was not a result
which activates GABAergic intercalated cells (ITC) within the amygdala. ITCinhibition of the Ce competes with BLA excitation of Ce, effectively cancel-
of tone stimulation or acquisition of fear conditioning. c-Fos is a
ing fear responses. Potentiation of PFC responses to the CS and inhibition of
marker of cellular activity but also can act as a transcription factor
conditioned fear responses also may involve reciprocal PFC interactions
with hippocampal, thalamic, and neocortical pathways. Modified with per-
are consistent with a role of gene expression in extinction
increased c-Fos expression in amygdala ITC cells
the spontaneous recovery of conditioned freezing that normally
ITC cells also exhibit NMDA-mediated plasticity
is observed with the passage of time.
suggesting that they may participate in long-term
An additional approach to enhancing mPFC function is the
use of metabolic enhancers such as methylene blue (MB), whichimprove activity-dependent brain energy production by targeting
Conflicting Lesion Evidence on the Role of the mPFC in Extinction
memory-improving action of MB in rats first was demonstratedfor inhibitory avoidance learning
Although there is much physiological evidence in favor of a
functional role of mPFC in learning and expression of extinction,
administration of MB could enhance retention of an extinguished
there also are conflicting lesion reports. Two groups did not find
conditioned response. Postextinction freezing was 50% lower in
any effect of pretraining mPFC lesions on extinction of condi-
rats that were receiving 4 mg/kg of MB, a dose that chronically is
used in human beings without negative side effects
another study found that lesions made after conditioning did not
Control rats injected with MB showed no changes in motor
impair subsequent extinction Interpretation
activity or general fearfulness, suggesting that postextinction MB
of permanent lesion effects often is hampered by potential
administration specifically enhanced memory for extinction. Rats
recovery of function or compensation by other structures. There
with improved retention of extinction also showed a greater
is a pressing need, therefore, for studies that use temporary
relative increase in cytochrome oxidase activity in the same
inactivation of mPFC via microinfusion of local anesthetics or the
prefrontal cortical regions that are activated during extinction
GABA antagonist muscimol. Preliminary reports using these
techniques are conflicting, showing increased fear
augmenting extinction-induced potentiation of mPFC. Note the
parallel with electrical stimulation and unit-recording findings
Conversely, decreases in cytochrome oxidase activity
challenge for future studies will be to identify the factors that
in the prefrontal cortex produced by genetic selection of rats that
could account for variability between laboratories. These might
are predisposed to helplessness results in
include contextual variables (e.g., AAA vs. ABB designs), the
rats with deficits in fear extinction that simulate the PTSD
presence of a competing appetitive instrumental response (such
as bar-pressing for food), or the number of extinction trials (e.g.,overtraining-induced masking of effects). Another possible rea-son for negative lesion effects is that the mPFC likely is part of a
Relevance to Treatment of Psychiatric Disorders
network of structures that collectively consolidate and expressextinction memory Disconnection of a
There is great interest in finding more effective treatments for
sufficient number of structures within the network may be a
anxiety disorders, which are among the most common mental
prerequisite for observing lesion deficits. Finally, recent studies
health problems. Extinction deficits have been implicated as a
show that mPFC neurons can signal acquisition of fear condi-
possible risk factor for the development of PTSD
show reduced extinction of aversively conditioned responses
separate modules within mPFC for exciting versus inhibiting fear.
and show impairments in afunctional network involving the amygdala and anterior cingu-
Enhancing Prefrontal Function Strengthens Extinction
late Brain-imaging studies ofPTSD patients show reduced activity
If prefrontal activation is essential for extinction learning, then
stimulating prefrontal cortex should strengthen extinction. Support
prefrontal cortex, an area that is homologous with extinction-
for this idea comes from experiments using electrical stimulation
related regions of rodent mPFC These studies
and metabolic enhancers. Electrical stimulation was used to mimic
also show increased amygdala activity in PTSD patients who are
short-latency tone-evoked responses of infralimbic neurons (100 –
suggesting a lack of top-down control of the amygdala by
Pairing this brief IL stimulation with conditioned tones reduced the
structures involved in extinction of fear.
expression of freezing, consistent with feed-forward inhibition of
Several recent functional imaging and volumetric studies dem-
amygdala output neurons mPFC stimulation also
onstrate that extinction activates perigenual and associated regions
strengthened extinction learning as evidenced by persistent de-
creased fear responses the day after the stimulation, suggesting LTP
of extinction-related synapses in mPFC.
coworkers demonstrated that retention of fear extinction was cor-
The role of LTP was tested directly by enhancing mPFC
related with the thickness of the vmPFC suggest-
responsiveness to MD thalamic inputs by applying high-fre-
ing that the likelihood of developing PTSD depends on the integrity
quency stimulation before extinction training
of the prefrontal extinction system. For a complete review of the
MD stimulation had no effect on the rate of extinction
learning within the training session, supporting lesion and
Behavioral therapy for PTSD (exposure therapy) is based
unit-recording findings that mPFC is not responsible for short-
term extinction memory. One week later, however, retention of
Therefore, methods of facilitating extinction and preventing the
extinction was markedly improved in potentiated rats, as evi-
return of fear may lead to more effective therapeutic interven-
denced by low rates of spontaneous recovery of freezing.
tions. Current behavioral techniques such as flooding and implo-
Improvement in extinction retention was correlated with poten-
sion could be improved by pharmacological interventions that
tiation of mPFC evoked potentials. Thus, mPFC LTP prevented
accelerate and strengthen extinction. For example, a reduction in
the number of exposure sessions required to successfully extin-
Berretta S, Pantazopoulos H, Caldera M, Pantazopoulos P, Paré D (2005):
guish fear responses could counteract the relatively high dropout
Infralimbic cortex activation increases c-Fos expression in intercalated
rate that is observed with this type of therapy
neurons of the amygdala. Neuroscience 132:943–953.
Bouton ME (2002): Context, ambiguity, and unlearning: Sources of relapse
after behavioral extinction. Biol Psychiatry 52:976 –986.
Several recent studies have shown that extinction learning in
Bremner JD (2002): Neuroimaging studies in post-traumatic stress disorder.
rats can be accelerated and strengthened with systemically
Curr Psychiatry Rep 4:254 –263.
applied drugs. These include the noradrenergic antagonist yo-
Bremner JD (2003): Functional neuroanatomical correlates of traumatic
himbine the dopamine D2 receptor antagonist
stress revisited 7 years later, this time with data. Psychopharmacol Bull
Burgos-Robles A, Santini E, Quirk GJ (2004): Blockade of NMDA receptors in
the medial prefrontal cortex impairs consolidation of fear extinction. SocNeurosci Abstr Program No. 328.14.
With the exception of DCS, the locus of action
Butter CM, Mishkin M, Rosvold HE (1963): Conditioning and extinction of a
in the brain of these drugs is not yet known. On the basis of
food-rewarded response after selective ablation of frontal cortex in rhe-
previous work, however, modulation of dopaminergic and nor-
sus monkeys. Exp Neurol 7:65–75.
adrenergic systems in the rat mPFC is likely to modulate the rate
Cain CK, Blouin AM, Barad M (2004): Adrenergic transmission facilitates
extinction of conditional fear in mice. Learn Mem 11:179 –187.
DCS has been shown to be effective when infused into the
Callaway NL, Riha PD, Wrubel KM, McCollum D, Gonzalez-Lima F (2002):
BLA and clinical studies show that adminis-
Methylene blue restores spatial memory retention impaired by an inhib-itor of cytochrome oxidase in rats. Neurosci Lett 332:83– 86.
tering DCS to acrophobic subjects who are undergoing exposure
Cassell MD, Wright DJ (1986): Topography of projections from the medial
therapy improves the effectiveness of the therapy
prefrontal cortex to the amygdala in the rat. Brain Res Bull 17:321–333.
However, there are some limitations to this approach, such
Chan KH, Morell JR, Jarrard LE, Davidson TL (2001): Reconsideration of the
as CS nonspecificity and tolerance to repeated DCS
role of the hippocampus in learned inhibition. Behav Brain Res 119:111–
the metabolic enhancer MB A metabolic approach
Charney DS (2004): Psychobiological mechanisms of resilience and vulner-
differs from the transmitter-receptor approach because it is not
ability: Implications for successful adaptation to extreme stress. AmJ Psychiatry 161:195–216.
selective for a single transmitter system or brain region but targets all
Charney DS, Deutch AY, Krystal JH, Southwick SM, Davis M (1993): Psycho-
the synapses that require increased energy during postextinction
biologic mechanisms of posttraumatic stress disorder. Arch Gen Psychi-
memory consolidation, such as in the various prefrontal cortex
Chhatwal JP, Davis M, Maguschak KA, Ressler KJ (2005): Enhancing cannabi-
activating mPFC during exposure therapy could include repetitive
noid neurotransmission augments the extinction of conditioned fear.
transcranial magnetic stimulation deep brain
Neuropsychopharmacology 30:516 –524.
Cohen H, Kaplan Z, Kotler M, Kouperman I, Moisa R, Grisaru N (2004): Repet-
itive transcranial magnetic stimulation of the right dorsolateral prefron-
Thus, prefrontal activation achieved pharmacologically,
tal cortex in posttraumatic stress disorder: A double-blind, placebo-
physiologically, or psychologically could serve as a useful adjunct to
controlled study. Am J Psychiatry 161:515–524.
exposure therapy by strengthening memory for the extinction
Condé F, Maire-Lepoivre E, Audinat E, Crepel F (1995): Afferent connections
of the medial frontal cortex of the rat. II. Cortical and subcortical affer-ents. J Comp Neurol 352:567–593.
Corcoran KA, Maren S (2003): Reversible inactivation of medial prefrontal
cortex disrupts the retrieval of fear extinction. Soc Neurosci Abstr 199.8. This work was supported by NIH grants: R01-MH58883,
Cox J, Westbrook RF (1994): The NMDA receptor antagonist MK-801 blocks
S06-GM08239, and R21-MH072156 (to GJQ), the Philippe Foun-
acquisition and extinction of conditioned hypoalgesic responses in the
dation (to RG), and NIH grant R01-NS37755 (to FGL).
rat. Q J Exp Psychol B 47:187–210. Aspects of this work were presented at the conference “Extinc-
Falls WA, Miserendino MJ, Davis M (1992): Extinction of fear-potentiated
tion: The Neural Mechanisms of Behavior Change”, February
startle: Blockade by infusion of an NMDA antagonist into the amygdala. 2– 6, 2005 in Ponce, Puerto Rico. The Conference was sponsored
Farinelli M, Deschaux O, Hugues S, Thevenet A, Garcia R (in press): Hip-
by the National Institute of Mental Health, National Institute on
pocampal train stimulation modulates recall of fear extinction indepen-
Drug Abuse, Ponce School of Medicine, University of Puerto Rico
dently of prefrontal cortex synaptic plasticity and lesions. Learn Mem. Centers of Biomedical Research Excellence (COBRE) Program,
Floyd NS, Price JL, Ferry AT, Keay KA, Bandler R (2001): Orbitomedial prefron-
Pfizer Global Pharmaceutical, and the Municipality of Ponce.
tal cortical projections to hypothalamus in the rat. J Comp Neurol 432:307–328.
Abelson JL, Curtis GC, Sagher O, Albucher RC, Harrigan M, Taylor SF, et al
Garcia R (2002): Stress, synaptic plasticity, and psychopathology. Rev Neuro-
(2005): Deep brain stimulation for refractory obsessive-compulsive dis-
order. Biol Psychiatry 57:510 –516.
Gewirtz JC, Falls WA, Davis M (1997): Normal conditioned inhibition and
Anderson P, Jacobs C, Rothbaum BO (2004): Computer-supported cognitive
extinction of freezing and fear-potentiated startle following electrolytic
behavioral treatment of anxiety disorders. J Clin Psychol 60:253–267.
lesions of medical prefrontal cortex in rats. Behav Neurosci 111:712–726.
Anglada-Figueroa D, Quirk GJ (2005): Lesions of the basal amygdala block
Gilboa A, Shalev AY, Laor L, Lester H, Louzoun Y, Chisin R, et al (2004):
expression of conditioned fear, but not extinction. J Neurosci 25:9680 –
Functional connectivity of the prefrontal cortex and the amygdala in
posttraumatic stress disorder. Biol Psychiatry 55:263–272.
Baeg EH, Kim YB, Jang J, Kim HT, Mook-Jung I, Jung MW (2001): Fast spiking
Gonzalez-Lima F, Bruchey AK (2004): Extinction memory improvement by
and regular spiking neural correlates of fear conditioning in the medial
the metabolic enhancer methylene blue. Learn Mem 11:633– 640.
prefrontal cortex of the rat. Cereb Cortex 11:441– 451.
Gottfried JA, Dolan RJ (2004): Human orbitofrontal cortex mediates extinc-
Baker JD, Azorlosa JL (1996): The NMDA antagonist MK-801 blocks the
tion learning while accessing conditioned representations of value. Nat
extinction of Pavlovian fear conditioning. Behav Neurosci 110:618 – 620.
Barrett D, Shumake J, Jones D, Gonzalez-Lima F (2003): Metabolic mapping
Hermans D, Dirikx T, Vansteenwegenin D, Baeyens F, Van den Bergh O, Eelen
of mouse brain activity after extinction of a conditioned emotional re-
P (2005): Reinstatement of fear responses in human aversive condition-
sponse. J Neurosci 23:5740 –5749.
ing. Behav Res Ther 43:533–551.
Herry C, Garcia R (2002): Prefrontal cortex long-term potentiation, but not
McDonald AJ, Mascagni F, Guo L (1996): Projections of the medial and lateral
long-term depression, is associated with the maintenance of extinction
prefrontal cortices to the amygdala: A Phaseolus vulgaris leucoaggluti-
of learned fear in mice. J Neurosci 22:577–583.
nin study in the rat. Neuroscience 71:55–75.
Herry C, Garcia R (2003): Behavioral and paired-pulse facilitation analyses of
Mickley GA, Kenmuir CL, Yocom AM, Wellman JA, Biada JM (2005): A role for
long-lasting depression at excitatory synapses in the medial prefrontal
prefrontal cortex in the extinction of a conditioned taste aversion. Brain
cortex in mice. Behav Brain Res 146:89 –96.
Herry C, Vouimba RM, Garcia R (1999): Plasticity in the mediodorsal thalamo-
Milad MR, Quinn BT, Pitman RK, Orr SP, Fischl B, Rauch SL (2005): Thickness of
prefrontal cortical transmission in behaving mice. J Neurophysiol 82:
ventromedial prefrontal cortex in humans is correlated with extinction
memory. Proc Natl Acad Sci U S A 102:10706 –10711.
Hugues S, Chessel A, Léna I, Marsault R, Garcia R (in press): Prefrontal infusion
Milad MR, Quirk GJ (2002): Neurons in medial prefrontal cortex signal mem-
of PD098059 immediately after fear extinction training blocks extinc-
ory for fear extinction. Nature 420:70 –74.
tion-associated prefrontal synaptic plasticity and decreases prefrontal
Milad MR, Rauch SL, Pitman RK, Quirk GJ (in press): Fear extinction in rats:
ERK2 phosphorylation. Synapse.
Implications for human brain imaging and anxiety disorders. Biol
Hugues S, Deschaux O, Garcia R (2004): Postextinction infusion of a mitogen-
activated protein kinase inhibitor into the medial prefrontal cortex im-
Milad MR, Vidal-Gonzalez I, Quirk GJ (2004): Electrical stimulation of medial
pairs memory of the extinction of conditioned fear. Learn Mem 11:540 –
prefrontal cortex reduces conditioned fear in a temporally specific man-
ner. Behav Neurosci 118:389 –394.
Hurley KM, Herbert H, Moga MM, Saper CB (1991): Efferent projections of the
Morgan MA, LeDoux JE (1995): Differential contribution of dorsal and ventral
infralimbic cortex of the rat. J Comp Neurol 308:249 –276.
medial prefrontal cortex to the acquisition and extinction of conditioned
Jay TM, Burette F, Laroche S (1995): NMDA receptor-dependent long-term
fear in rats. Behav Neurosci 109:681– 688.
potentiation in the hippocampal afferent fibre system to the prefrontal
Morgan MA, Romanski LM, LeDoux JE (1993): Extinction of emotional learn-
cortex in the rat. Eur J Neurosci 7:247–250.
ing: Contribution of medial prefrontal cortex. Neurosci Lett 163:109 –113.
Jay TM, Witter MP (1991): Distribution of hippocampal CA1 and subicular
Morgan MA, Schulkin J, LeDoux JE (2003): Ventral medial prefrontal cortex
efferents in the prefrontal cortex of the rat studied by means of antero-
and emotional perseveration: The memory for prior extinction training.
grade transport of Phaseolus vulgaris-leucoagglutinin. J Comp NeurolBehav Brain Res 146:121–130.
Morrow BA, Elsworth JD, Rasmusson AM, Roth RH (1999): The role of meso-
Kaczmarek L (2002): c-Fos in learning: Beyond the mapping of neuronal
prefrontal dopamine neurons in the acquisition and expression of con-
activity. In: Kaczmarek L, Robertson HA, editors. Handbook of Chemical
ditioned fear in the rat. Neuroscience 92:553–564. Neuroanatomy: Immediate Early Genes and Inducible Transcription Factors
Myers KM, Davis M (2002): Behavioral and neural analysis of extinction. in Mapping of the Central Nervous System Function and Dysfunction. Am-
Myers KM, Davis M (2004): Temporary inactivation of the infralimbic region
Kandel ER (2001): The molecular biology of memory storage: A dialogue
of medial prefrontal cortex via localized infusion of muscimol or ba-
between genes and synapses. Science 294:1030 –1038.
clofen does not impair conditioned inhibition or extinction of fear po-
Kimble DP, Kimble RJ (1970): The effect of hippocampal lesions on extinction
tentiated startle in rats. Soc Neurosci Abstr 328.2.
and “hypothesis” behavior in rats. Physiol Behav 5:735–738.
Naylor GJ, Martin B, Hopwood SE, Watson Y (1986): A two-year double-blind
Konorski J (1967): Integrative Activity of the Brain. Chicago: University of
crossover trial of the prophylactic effect of methylene blue in manic-
depressive psychosis. Biol Psychiatry 21:915–920.
Laviolette SR, Lipski WJ, Grace AA (2005): A subpopulation of neurons in the
Paré D, Smith Y (1993a): Distribution of GABA immunoreactivity in the
medial prefrontal cortex encodes emotional learning with burst and
amygdaloid complex of the cat. Neuroscience 57:1061–1076.
frequency codes through a dopamine D4 receptor-dependent basolat-
Paré D, Smith Y (1993b): The intercalated cell masses project to the central
eral amygdala input. J Neurosci 25:6066 – 6075.
and medial nuclei of the amygdala in cats. Neuroscience 57:1077–1090.
Lazar SW, Bush G, Gollub RL, Fricchione GL, Khalsa G, Benson H (2000):
Parnas AS, Weber M, Richardson R (2005): Effects of multiple exposures to
Functional brain mapping of the relaxation response and meditation. Neuroreport 11:1581–1585.
D-cycloserine on extinction of conditioned fear in rats. Neurobiol Learn
Lebron K, Milad MR, Quirk GJ (2004): Delayed recall of fear extinction in rats
with lesions of ventral medial prefrontal cortex. Learn Mem 11:544 –548.
Pavlov I (1927): Conditioned Reflexes. London: Oxford University Press.
Ledgerwood L, Richardson R, Cranney J (2003): Effects of D-cycloserine on
Peri T, Ben Shakhar G, Orr SP, Shalev AY (2000): Psychophysiologic assess-
extinction of conditioned freezing. Behav Neurosci 117:341–349.
ment of aversive conditioning in posttraumatic stress disorder. Biol Psy-
Ledgerwood L, Richardson R, Cranney J (2005): D-cycloserine facilitates
extinction of learned fear: Effects on reacquisition and generalized ex-
Phelps EA, Delgado MR, Nearing KI, LeDoux JE (2004): Extinction learning in
tinction. Biol Psychiatry 57:841– 847.
humans: Role of the amygdala and vmPFC. Neuron 43:897–905.
Likhtik E, Pelletier JG, Paz R, Paré D (2005): Prefrontal control of the amyg-
Pirot S, Jay TM, Glowinski J, Thierry AM (1994): Anatomical and electrophys-
dala. J Neurosci 25:7429 –7437.
iological evidence for an excitatory amino acid pathway from the tha-
Lin CH, Yeh SH, Lu HY, Gean PW (2003): The similarities and diversities of
lamic mediodorsal nucleus to the prefrontal cortex in the rat. Eur J Neu-
signal pathways leading to consolidation of conditioning and consoli-
dation of extinction of fear memory. J Neurosci 23:8310 – 8317.
Ponnusamy R, Nissim HA, Barad M (2005): Systemic blockade of D2-like
Lissek S, Powers AS, McClure EB, Phelps EA, Woldehawariat G, Grillon C, et al
dopamine receptors facilitates extinction of conditioned fear in mice.
(2005): Classical fear conditioning in the anxiety disorders: A meta-anal-
ysis. Behav Res Ther 43:1391–1424.
Quirk GJ (2002): Memory for extinction of conditioned fear is long-lasting
Lu KT, Walker DL, Davis M (2001): Mitogen-activated protein kinase cascade
and persists following spontaneous recovery. Learn Mem 9:402– 407.
in the basolateral nucleus of amygdala is involved in extinction of fear-
Quirk GJ, Likhtik E, Pelletier JG, Paré D (2003): Stimulation of medial prefron-
potentiated startle. J Neurosci 21:RC162.
tal cortex decreases the responsiveness of central amygdala output
Maroun M, Richter-Levin G (2003): Exposure to acute stress blocks the induc-
neurons. J Neurosci 23:8800 – 8807.
tion of long-term potentiation of the amygdala-prefrontal cortex path-
Quirk GJ, Russo GK, Barron JL, Lebron K (2000): The role of ventromedial
way in vivo. J Neurosci 23:4406 – 4409.
prefrontal cortex in the recovery of extinguished fear. J Neurosci 20:
Martinez JL Jr, Jensen RA, Vasquez BJ, McGuinness T, McGaugh JL (1978):
Methylene blue alters retention of inhibitory avoidance responses.
Rauch SL, Shin LM, Phelps EA (2006): Neurocircuitry models of posttraumatic
stress disorder and extinction: human neuroimaging research past,
McCormick DA, Thompson RF (1982): Locus coeruleus lesions and resistance
present and future. Biol Psychiatry 60:376 –382.
to extinction of a classically conditioned response: Involvement of the
Rauch SL, Shin LM, Segal E, Pitman RK, Carson MA, McMullin K, et al (2003):
neocortex and hippocampus. Brain Res 245:239 –249.
Selectively reduced regional cortical volumes in post-traumatic stress
McDonald AJ (1991): Organization of amygdaloid projections to the pre-
disorder. Neuroreport 14:913–916.
frontal cortex and associated striatum in the rat. Neuroscience 44:1–14.
Rescorla RA (2004): Spontaneous recovery. Learn Mem 11:501–509.
Rescorla RA, Heth CD (1975): Reinstatement of fear to an extinguished
Sierra-Mercado D, Burgos-Robles A, Corcoran KA, Lebron K, Quirk GJ (2005):
conditioned stimulus. J Exp Psychol Anim Behav Process 1:88 –96.
Inactivation of the ventromedial prefrontal cortex reduces the expres-
Ressler KJ, Rothbaum BO, Tannenbaum L, Anderson P, Graap K, Zimand E, et
sion of conditioned fear. Soc Neurosci Abstr 649.9.
al (2004): Cognitive enhancers as adjuncts to psychotherapy: Use of
Sokoloff L (1992): Imaging techniques in studies of neural functions. In:
D-cycloserine in phobic individuals to facilitate extinction of fear. Arch
Gonzalez-Lima F, Finkenstadt T, Scheich H, editors. Advances in MetabolicGen Psychiatry 61:1136 –1144. Mapping Techniques for Brain Imaging of Behavior and Learning Functions.
Rhodes SE, Killcross S (2004): Lesions of rat infralimbic cortex enhance recov-
ery and reinstatement of an appetitive Pavlovian response. Learn Mem
Song E, Kim JJ (2004): Effects of auditory cortex lesions on acquisition and
extinction of fear conditioning to two different auditory stimuli in rats.
Riha PD, Bruchey AK, Echevarria DJ, Gonzalez-Lima F (2005): Memory facili-
tation by methylene blue: Dose-dependent effect on behavior and brain
Sotres-Bayon F, Bush DE, LeDoux JE (2004): Emotional perseveration: An
oxygen consumption. Eur J Pharmacol 511:151–158.
update on prefrontal-amygdala interactions in fear extinction. Learn
Royer S, Martina M, Paré D (1999): An inhibitory interface gates impulse
traffic between the input and output stations of the amygdala. J Neurosci
Sotres-Bayon F, Cain CK, LeDoux JE (2006): Brain mechanisms of fear extinc-
tion: Historical perspectives on the contribution of prefrontal cortex. Biol
Royer S, Paré D (2002): Bidirectional synaptic plasticity in intercalated amyg-
dala neurons and the extinction of conditioned fear responses. Neuro-
Teich AH, McCabe PM, Gentile CC, Schneiderman LS, Winters RW, Liskowsky
DR, et al (1989): Auditory cortex lesions prevent the extinction of Pavlov-
Santini E, Ge H, Ren K, Pena DO, Quirk GJ (2004): Consolidation of fear
ian differential heart rate conditioning to tonal stimuli in rabbits. Brain
extinction requires protein synthesis in the medial prefrontal cortex.
van Minnen A, Hagenaars M (2002): Fear activation and habituation patterns
Santini E, Muller RU, Quirk GJ (2001): Consolidation of extinction learning
as early process predictors of response to prolonged exposure treat-
involves transfer from NMDA-independent to NMDA-dependent mem-ory. J Neurosci 21:9009 –9017.
ment in PTSD. J Trauma Stress 15:359 –367.
Shin LM, Orr SP, Carson MA, Rauch SL, Macklin ML, Lasko NB, et al (2004):
Vertes RP (2004): Differential projections of the infralimbic and prelimbic
Regional cerebral blood flow in the amygdala and medial prefrontal
cortex in the rat. Synapse 51:32–58.
cortex during traumatic imagery in male and female Vietnam veterans
Vouimba RM, Garcia R, Baudry M, Thompson RF (2000): Potentiation of condi-
with PTSD. Arch Gen Psychiatry 61:168 –176.
tioned freezing following dorsomedial prefrontal cortex lesions does not
Shin LM, Whalen PJ, Pitman RK, Bush G, Macklin ML, Lasko NB, et al (2001): An
interfere with fear reduction in mice. Behav Neurosci 114:720 –724.
fMRI study of anterior cingulate function in posttraumatic stress disor-
Walker DL, Davis M (2002): The role of amygdala glutamate receptors in fear
der. Biol Psychiatry 50:932–942.
learning, fear-potentiated startle, and extinction. Pharmacol Biochem
Shumake J, Barrett D, Gonzalez-Lima F (2005): Behavioral characteristics of
rats predisposed to learned helplessness: Reduced reward sensitivity,
Walker DL, Ressler KJ, Lu KT, Davis M (2002): Facilitation of conditioned fear extinc-
increased novelty seeking, and persistent fear memories. Behav Brain Res
tion by systemic administration or intra-amygdala infusions of D-cycloserine
as assessed with fear-potentiated startle in rats. J Neurosci 22:2343–2351.
Shumake J, Poremba A, Edwards E, Gonzalez-Lima F (2000): Congenital
Weible AP, McEchron MD, Disterhoft JF (2000): Cortical involvement in ac-
helpless rats as a genetic model for cortex metabolism in depression.
quisition and extinction of trace eyeblink conditioning. Behav Neurosci
Trastorno por déficit de atención/hiperactividad:estado de la cuestión y futuras líneas de investigación E. Cardo a, M. Servera b TRASTORNO POR DÉFICIT DE ATENCIÓN/HIPERACTIVIDAD: ESTADO DE LA CUESTIÓN Y FUTURAS LÍNEAS DE INVESTIGACIÓN Resumen. Objetivo . El presente trabajo es una revisión sobre el estado de la cuestión del trastorno por déficit de atención/hi- peractivi
Protocole « Epidémiologie de la prophylaxie secondaire des mycoses invasives » du Groupe de Travail des Maladies Infectieuses de la Société Allemande d’Hémato-Oncologie Introduction Prophylaxie primaire. La prophylaxie primaire antifongique des patients hémato-oncologiques a été examiné dans plusieures études1. Une réduction d’infection et de mortalité a été
 behavioral networks. Metabolic responses to a test tone were
compared in groups of mice that received fear conditioning, a
pseudorandom treatment (unpaired tones and shocks), or con-
ditioning followed by extinction. Consistent with single-unit andevoked-potential recording, the largest increase in metabolic activityafter extinction occurred in the mPFC. The infralimbic (but not theprelimbic) area showed significantly more metabolic activity thancontrols. In addition to IL, significant metabolic increases were
Conditioning Extinction
behavioral networks. Metabolic responses to a test tone were
compared in groups of mice that received fear conditioning, a
pseudorandom treatment (unpaired tones and shocks), or con-
ditioning followed by extinction. Consistent with single-unit andevoked-potential recording, the largest increase in metabolic activityafter extinction occurred in the mPFC. The infralimbic (but not theprelimbic) area showed significantly more metabolic activity thancontrols. In addition to IL, significant metabolic increases were
Conditioning Extinction