Ordered, immediately called back and the same day delivered the order.Very pleased with the work. Thank you for prompt and accurate work https://africarx.co.za/buy-levitra-south-africa.html great prices, delivered on the day of the order. Pleasant managers consult by phone.
Cond_106_311.567_579.tp
᭧ The Cooper Ornithological Society 2004
FACTORS RELATED TO FECAL ESTROGENS AND FECAL
BRIAN E. WASHBURN1,3, DOUGLAS J. TEMPEL2, JOSHUA J. MILLSPAUGH1,4, R. J. GUTIE´RREZ2
1Department of Fisheries and Wildlife Sciences, University of Missouri, 302 A-BNR Building,2Department of Fisheries, Wildlife, and Conservation Biology, University of Minnesota, St. Paul, MN 55108
We estimated concentrations of fecal reproductive steroid metabolites in free-
ranging California Spotted Owls (Strix occidentalis occidentalis) during the breeding season. We collected fresh fecal samples (n ϭ 142) from 65 individual owls in the Sierra Nevadaduring April–August of 2001. We developed and validated radioimmunoassay proceduresto quantify fecal estrogen metabolites and fecal testosterone metabolites. We used an infor-mation-theoretic approach to identify factors that might influence fecal estrogen (E), fecaltestosterone (T), and fecal estrogen:testosterone ratio (E:T ratio) levels during the owl’sbreeding season. We hypothesized that factors related to sampling procedures, owl charac-teristics (sex, reproductive status), and habitat might influence fecal reproductive steroidlevels. Our analyses suggested that sampling factors and owl characteristics, but not habitatvariables, were related to fecal reproductive steroid levels in Spotted Owls. Our most sup-ported models explained Ͻ30% of the observed variation. Fecal testosterone levels werehigher in male Spotted Owls than females, whereas E:T ratios were higher in femalescompared to males. High fecal estrogens were correlated with high fecal glucocorticoids innonbreeding Spotted Owls, whereas fecal estrogens and fecal glucocorticoids were not re-lated in breeding birds. Sampling influenced fecal reproductive steroid measures, and biasfrom small-mass fecal samples might partially explain these relationships. Noninvasive mea-surements of fecal reproductive steroids might be useful for sex determination and repro-ductive assessment of free-ranging Spotted Owls. However, more research is needed tounderstand the variability we observed in sex steroids before this technique can be effectivein conservation studies. California Spotted Owl, estrogens, glucocorticoids, noninvasive, testoster-one, physiology, Strix occidentalis occidentalis.
Factores Asociados con los Estro´genos Fecales y la Testosterona Fecal en
Estimamos las concentraciones de metabolitos esteroides reproductivos en
individuos silvestres de la especie Strix occidentalis occidentalis durante la e´poca repro-ductiva. Colectamos muestras fecales frescas (n ϭ 142) pertenecientes a 65 lechuzas en laSierra Nevada entre abril y agosto de 2001. Desarrollamos y validamos un procedimientode radioinmunoensayo para cuantificar metabolitos de estro´geno fecales y metabolitos detestosterona fecales. Utilizamos un enfoque informativo-teo´rico para identificar los factoresque podrı´an influenciar los niveles de estro´genos fecales (E), testosterona fecal (T) y elcociente entre estro´genos y testosterona (cociente E:T) durante la e´poca reproductiva de laslechuzas. Hipotetizamos que factores relacionados con los procedimientos de muestreo, ca-racterı´sticas de la lechuza (sexo, estado reproductivo) y el ha´bitat podrı´an influenciar losniveles de esteroides reproductivos en las fecas. Nuestros ana´lisis sugieren que los factoresasociados al muestreo y las caracterı´sticas de la lechuza se correlacionaron con los nivelesde esteroides reproductivos en las fecas, pero e´stos no se relacionaron con las variables deha´bitat. Nuestro modelo ma´s robusto explico´ Ͻ30% de la variacio´n observada. Los nivelesde testosterona fecal fueron mayores en los machos que en las hembras, mientras que elcuociente E:T fue mayor en las hembras que en los machos. En lechuzas no reproductivas,los niveles altos de estro´genos fecales se correlacionaron con niveles altos de glucocorti-coides fecales, mientras que en individuos reproductivos los estro´genos fecales y los glu-
Manuscript received 12 May 2003; accepted 27 March 2004. 3 Present address: USDA, Wildlife Services, National Wildlife Research Center, 6100 Columbus Avenue,
4 Corresponding author. E-mail: millspaughj@missouri.edu
corticoides fecales no se correlacionaron. Estas relaciones pueden ser explicadas en partepor la influencia del muestreo sobre las medidas de esteroides reproductivos fecales y porel sesgo causado por muestras fecales muy livianas. Los niveles de esteroides reproductivosfecales medidos con te´cnicas no invasivas pueden ser u´tiles para la determinacio´n de sexosy la evaluacio´n del estado reproductivo de individuos silvestres de la especie S. occidentalisoccidentalis. Sin embargo, antes de que esta te´cnica pueda ser efectiva en estudios deconservacio´n, se requiere de ma´s investigacio´n para entender la variabilidad que observamosen los niveles de esteroides sexuales.
Reproductive steroid hormone levels provide
important information regarding the reproduc-
The Northern and Mexican Spotted Owls (Strix
tive status of animals. However, longitudinal
occidentalis caurina and S. o. lucida) are listed
studies of reproductive function in free-ranging
in the United States as threatened subspecies,
Spotted Owls using traditional techniques (i.e.,
whereas the California subspecies (S. o. occi-
blood sampling) would require repeatedly sub-
dentalis) is not (USDI 1990, 1993, 2003). Be-
jecting owls to the stress of capture, restraint,
cause the conservation status of Spotted Owls
and blood collection (Le Maho et al. 1992). In
has great economic ramifications, there has been
contrast, noninvasive monitoring of reproductive
extensive research on its biology (Gutie´rrez et
steroids using fecal hormone metabolites is ad-
al. 1995, Franklin et al. 2004). The biological
vantageous because sample collection does not
status of the California Spotted Owl is of interest
require animal capture, samples can be collected
because forest management practices and habitat
without disturbing study animals, and numerous
conditions within its range are diverse (Verner
samples can be collected from individuals (Ber-
et al. 1992). In particular, Sierra Nevada forests,
covitz et al. 1982, Kofuji et al. 1993). Our ob-
where most California Spotted Owls reside, have
jectives were: (1) to validate radioimmunoassay
been managed using a wide variety of silvicul-
(RIA) procedures to quantify estrogen and tes-
tural practices, with much less emphasis on
tosterone metabolites in Spotted Owl feces and
clearcutting than in the range of the Northern
(2) to evaluate factors that might explain varia-
Spotted Owl. Also, the California Spotted Owl
tion in fecal estrogen (E), fecal testosterone (T),
is generally well distributed within its limited
and estrogen:testosterone ratio (E:T ratio) levels
in free-ranging California Spotted Owls during
Central to current Spotted Owl research has
been a desire to understand the mechanisms thatinfluence its population dynamics and habitat se-
lection (Franklin et al. 2000, 2004). Because ex-periments with this species are difficult to exe-
cute (Noon and Franklin 2002), alternative ap-
Our 925-km2 study area was located in the
proaches are being used to elucidate basic eco-
north-central Sierra Nevada between George-
town and South Lake Tahoe, California (39ЊN,
meta-analysis (Burnham et al. 1996), population
120ЊW). During 1986–2001, we surveyed a core
genetics (Barrowclough et al. 1999), and the ef-
area of 355 km2 each year for all Spotted Owls
fects of disturbance using noninvasive tech-
plus selected territories within an adjacent 570
niques (Wasser et al. 1997, Tempel and Gutie´r-
km2 (Seamans et al. 2001). Public (USDA Forest
rez 2004). A field protocol exists for assessing
Service) and private land comprised 63% and
reproductive status in owls (Franklin et al.
1996), but a negative assessment using this pro-
Elevation on the study area ranged from 366
tocol does not always mean that the bird did not
m to 2257 m. Vegetation was typical Sierran
attempt to nest. One noninvasive technique that
mixed-conifer forest (Ku¨chler 1977, Rundel et
might provide useful insight into the reproduc-
al. 1977). From 600 m to 1500 m the forests
tive biology of the Spotted Owl is the evaluation
were dominated by ponderosa pine (Pinus pon-
of fecal reproductive hormone levels (i.e., tes-
derosa) on more xeric sites and white fir (Abies
tosterone and estrogen) as a measure of physi-
concolor) on more mesic sites. Above 1500 m
ological response to social and environmental
there was a transition zone dominated by red fir
(Abies magnifica). Other common tree species
that occurred within the study area included sug-
lowed the ICN protocol for the total estrogens
ar pine (Pinus lambertiana), Douglas-fir (Pseu-
I125 RIA. We measured testosterone metabolites
dotsuga menziesii), incense cedar (Calocedrus
in Spotted Owl feces using a commercially
decurrens), canyon live oak (Quercus chrysole-
available testosterone I125 double-antibody radio-
pis), California black oak (Quercus kelloggii),
immunoassay kit (Cat. #DSL-4100, Diagnostic
and Pacific dogwood (Cornus nuttallii).
Systems Laboratories, Webster, Texas). We fol-lowed the manufacturer’s method for the testos-
terone I125 RIA, except that we halved the vol-
We collected fecal samples from April to August
ume of all reagents. We conducted standard val-
2001, with early April marking the onset of
idations including parallelism and recovery of
Spotted Owl nesting on the study area. We lo-
exogenous sex steroid validation assays (Jeff-
cated owls using standard techniques (Forsman
coate 1981, Grotjan and Keel 1996, O’Fegan
1983). We placed a clean polyethylene sheet be-
2000) on six pooled fecal extracts to confirm
neath a roosting owl (when possible) to facilitate
these assays were accurately and precisely mea-
sample collection. We removed the fecal portion
suring estrogen and testosterone metabolites in
of excreta from the urine portion (Wasser et al.
Spotted Owl feces. Pooled fecal extracts (ex-
1997) and placed the fecal samples into cryo-
pected low, medium, and high levels) were com-
genic vials. Fecal samples were then placed into
posed of fecal extracts from three males and
liquid nitrogen or placed on ice until they could
three females. We added exogenous testosterone
be transferred to liquid nitrogen. We collected
to the medium and high pool fecal extracts to
fecal samples from adults and did not use cecal
obtain testosterone values under higher dilution
discharge samples (Tempel and Gutie´rrez 2004).
levels. Interassay variation was calculated from
Most individual owls on the study area were col-
the three owl samples. Intra-assay variation was
or banded, and unmarked birds were captured
calculated by averaging the CVs of replicate
and color banded at the first opportunity. We
tubes from 20 randomly chosen samples. We
used these color bands to assign fecal samples
used tests for equal slopes (parallelism) to de-
to individuals. The sex of all birds was deter-
termine if log-transformed curves of serially di-
mined by vocal characteristics or nesting behav-
luted pool fecal extracts were parallel to log-
ior (Forsman 1983, Franklin et al. 1996).
transformed total estrogen and testosterone stan-dard curves (Neter et al. 1990). We used SAS
(SAS Institute 1985) to perform all statistical
We placed frozen fecal samples in a lyophilizer
(Freeze-dry Specialties, Inc., Osseo, Minnesota)for 24 hr. Once samples were freeze-dried, we
FACTORS RELATED TO REPRODUCTIVESTEROID LEVELS
ground them, sifted them through a stainlesssteel mesh to remove large particles, and mixed
Based on the literature, we developed a priori
them thoroughly. We weighed each fecal sample
verbal hypotheses regarding the correlation of
to the nearest 0.001 g. We extracted glucocor-
various factors with fecal estrogen, fecal testos-
ticoids from feces using a modification of
terone, and estrogen:testosterone (E:T) ratio lev-
Schwarzenberger et al. (1991). We placed dried
els in California Spotted Owls during the breed-
feces (ϳ0.1 g) in a test tube with 2.0 mL of 90%
ing season. First, we hypothesized that sample
methanol and vortexed at high speed in a mul-
collection or storage factors might influence fe-
titube vortexer for 30 min. Samples were then
cal reproductive steroid concentrations (sam-
centrifuged at 500 g for 20 min, and the super-
pling hypothesis). Second, we hypothesized that
natant was saved and stored atϪ84ЊC.
characteristics of individual owls and the timingof sample collection might influence fecal repro-
ductive steroid levels (biological hypothesis).
Last, we hypothesized that habitat characteristics
We quantified estrogen metabolite levels in
might be related to fecal reproductive steroid
Spotted Owl fecal extracts using a commercially
levels (habitat hypothesis). For each hypothesis,
available total estrogens I125 double-antibody ra-
we developed a set of a priori models. We used
dioimmunoassay kit (Cat. #07–140202, ICN
several covariates to develop the set of a priori
Biomedicals, Costa Mesa, California). We fol-
candidate models representing multiple hypoth-
eses of covariate effects on estrogen metabolites,
between territory centers (Peery et al. 1999,
testosterone metabolites, and E:T ratios in owl
Franklin et al. 2000). A territory center was the
location of a nest site, or for non-nesting owls,
the average location of roost sites recorded at
Sampling covariates. Sampling covariates in-
least 1 week apart. Most roost sites within a ter-
cluded the initial storage method for collected
ritory were near each other within the same for-
samples (SSTOR) and the mass of an individual
est stand. Mean nearest-neighbor distance was
fecal sample (SMASS). Storage methods includ-
estimated for the year of highest owl density on
ed placing vials containing feces into liquid ni-
trogen within 6 hr of collection or storage of
We defined owl habitat as all mature conifer
vials on ice for up to 36 hr before storage in
forest, irrespective of canopy cover, and medi-
liquid nitrogen. Sample mass was the mass of
um-sized conifer forest with canopy cover
the fecal sample (to the nearest 0.001 g) after
Ն70% because California Spotted Owls nest,
freeze-drying and sieving. Tempel and Gutie´rrez
roost, and forage in these habitats (Gutie´rrez et
(2004) found that fecal glucocorticoids were
al. 1992). Mature forest consisted of stands hav-
higher in Spotted Owl feces initially stored on
ing dominant trees Ն61 cm diameter at breast
ice and in samples of small mass. Thus, we pre-
height (dbh), and mid-seral forest had dominant
dicted fecal estrogens and fecal testosterone
trees ranging from 30 to 60 cm dbh. We esti-
would be higher in samples initially stored on
mated habitat variables as described in Tempel
ice and samples having smaller mass (Appendix
Franklin et al. (2000) found that territory-lev-
Biological covariates. Biological covariates
el habitat characteristics (e.g., the amount of
included sex of the bird (SEX), reproductive
core owl habitat within a Spotted Owl’s territo-
life-history stage (RLHS) of the bird, fecal glu-
ry) were important predictors of reproductive
cocorticoid level (CORT), and sample collection
output in Northern Spotted Owls. Thus, we hy-
date (DATE). We determined the sex of an in-
pothesized that the amount of core area in the
dividual owl from its unique color band or by
owl’s territory or the number of discrete habitat
the pitch of its call (Forsman et al. 1984). We
patches in the owl’s territory influenced repro-
hypothesized that male Spotted Owls would
ductive steroid levels in California Spotted
have higher fecal testosterone levels than fe-
Owls. Specifically, we predicted fecal estrogen
males, whereas females would have higher fecal
and fecal testosterone concentrations would be
estrogen and E:T ratios than males (Appendix
higher in owls having more core area and fewer
B). We estimated the reproductive life-history
habitat patches in their territory (Appendix B).
stage of each individual owl using the methods
Model selection. We used information-theo-
described in Franklin et al. (1996). Each indi-vidual owl was classified as nonbreeding or
retic model selection (Burnham and Anderson
breeding (i.e., actively nesting or fledging
young). We expected breeding owls to have
MIXED in SAS) where individual owls were the
higher fecal reproductive steroid levels than
sampling unit, the date of fecal sample collec-
nonbreeding owls. Glucocorticoid metabolite
tion was a repeated effect, and all other covaria-
concentrations were estimated as described in
tes were fixed effects (Littell et al. 1996). Po-
Tempel and Gutie´rrez (2004). Since elevated
tential nonindependence among fecal samples
corticosterone levels might suppress reproduc-
from the same individual was accounted for by
tion in birds (Wingfield 1988, Wingfield et al.
the repeated-measures analysis. To account for
1998), we hypothesized that owls with high fe-
possible heterogeneous sampling variances
cal glucocorticoid levels would have lower re-
among individuals, we first used restricted max-
imum-likelihood estimation to select the appro-
Habitat covariates. Habitat covariates includ-
priate covariance structure for the global model
ed the amount of core area in the owl’s territory
using PROC MIXED (Littell et al. 1996); the
(CORE) and the number of discrete habitat
sample variance structures were ranked using
patches in the owl’s territory (PATCH). We de-
AIC (a small-sample-size correction for Akai-
fined an owl territory as a circle with a radius
ke’s Information Criterion). The diagonal matrix
of one-half the mean nearest-neighbor distance
grouped by sex was the best fit for estrogens and
Likelihood-ratio tests for goodness-of-fit of subglobal models estimating fecal estrogens, fecal tes-
tosterone, and fecal estrogens:testosterone ratios in California Spotted Owls in the north-central Sierra Nevada,California, April–August 2001. Explanations of covariate abbreviations appear in Appendix A. A priori modelsthat contained covariates from subglobal models that were not significant were not included in the final model-selection subset.
a The Sampling subglobal model contained SMASS, SSTOR, and SMASS*SSTOR. b The SEX*RLHS subglobal model contained SEX, RLHS, and SEX*RLHS. c The CORT*RLHS subglobal model contained CORT, RLHS, and CORT*RLHS. d The CORT*DATE subglobal model contained CORT, DATE, (DATE)2, and CORT*DATE. e The habitat subglobal model contained CORE, PATCH, and CORE*PATCH.
testosterone, whereas the Gaussian spatial struc-
samples were diluted with steroid dilutent to 1:
ture grouped by sex was the best fit for E:T.
64 (e.g., 50 L of fecal extract into 1.55 mL of
We used a two-stage approach to model fitting
steroid dilutent) prior to assay. Intra-assay vari-
(Franklin et al. 2000, Seamans et al. 2001). First,
ation was 2.4%, calculated from 20 randomly
we compared models containing only sampling
chosen samples, and interassay variation for five
covariates. We used AIC to select the sampling
assays was 11.2%. The manufacturer’s reported
model with the most support for each of the
cross-reactivity of ICN total estrogens antisera
three hormone response variables. Second, we
was 100% with estrone and estradiol 17-, 9%
compared models containing biological and hab-
with estriol, 7% with estradiol 17-␣, and Ͻ1%
itat covariates. To each of the a priori biological
for other steroids. Assay sensitivity was 100 pg
and habitat models, we also included the cova-
riate(s) from the best-fit sampling model. We as-
Fecal testosterone. The DSL testosterone I125
sessed goodness-of-fit using likelihood-ratio
RIA reliably quantified testosterone metabolites
tests for a series of subglobal models and re-
in Spotted Owl feces. Serial dilutions (1:2 up to
moved those models which did not have signif-
1:128) of low, medium, and high pool fecal ex-
icant fit from consideration (Table 1). We con-
tracts yielded displacement curves that were par-
sidered test results significant for P Ͻ 0.05. For
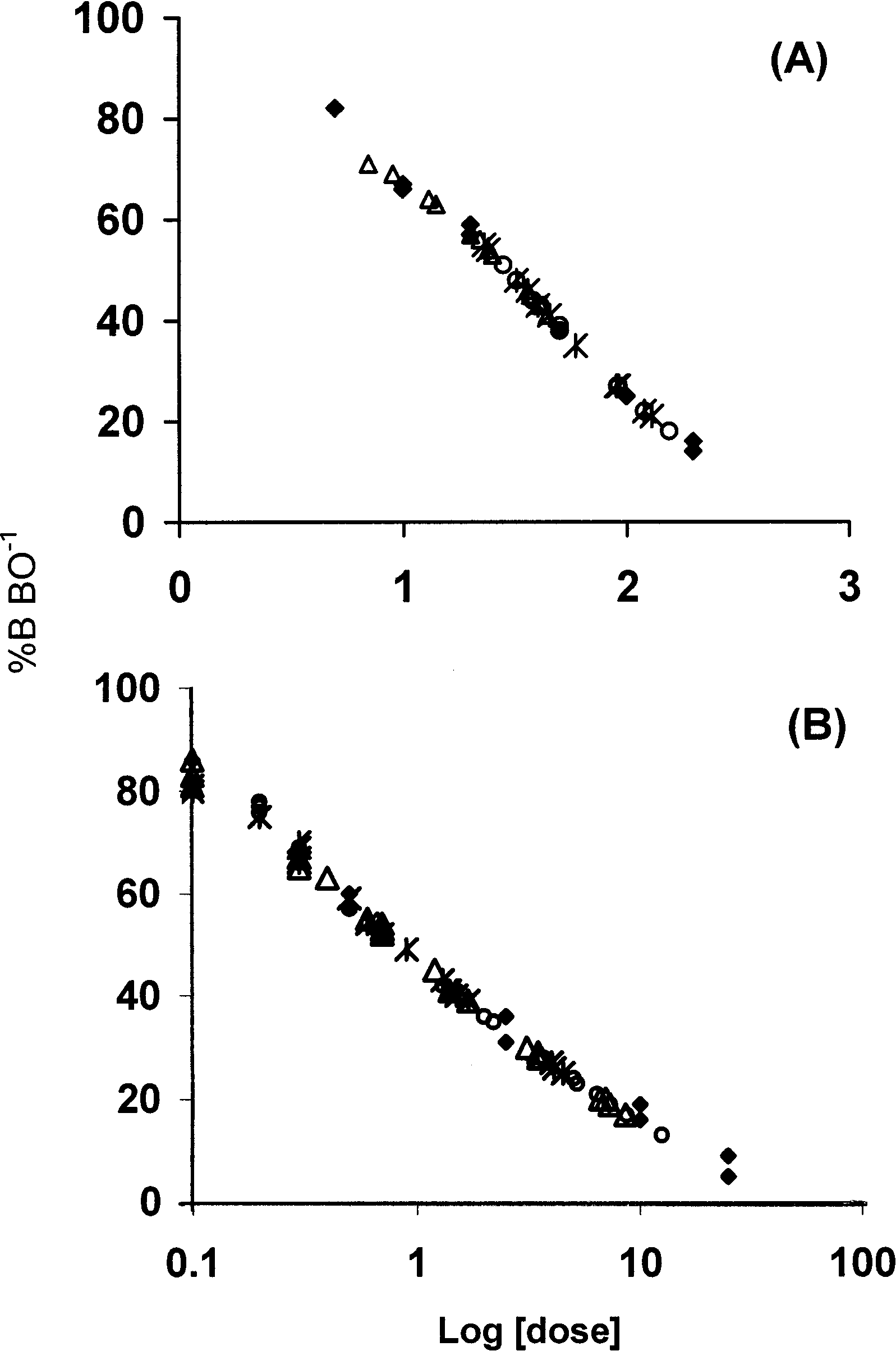
allel to the testosterone standard curve (Fig. 1;
the final evaluation, a priori models were ranked
all P Ͼ 0.3). Mean recovery of exogenous tes-
using AIC (Burnham and Anderson 1998).
tosterone (range 0.5–10 ng mLϪ1; levels chosen
to correspond with expected fecal testosterone
levels from actual samples) added to low and
high pool fecal extracts was 94.1 Ϯ 2.9% (n ϭ
Fecal estrogens. The I125 RIA reliably quantified
18). Acceptable recovery of exogenous analyte
estrogen metabolites in Spotted Owl feces. Se-
(within 90–110%) and demonstration of paral-
rial dilutions (1:2 up to 1:128) of low, medium,
lelism suggested no sample matrix effects (Jeff-
and high pool fecal extracts yielded displace-
coate 1981, Grotjan and Keel 1996, O’Fegan
ment curves parallel to the estrogens standard
2000). Extracts from fecal samples were not di-
curve (Fig. 1; all P Ͼ 0.2). Mean recovery of
luted in assay dilutent prior to assay. Intra-assay
exogenous total estrogens (range 10–50 pg
variation, calculated from 20 randomly chosen
mLϪ1) added to low and high pool fecal extracts
samples, was 2.2%. Interassay variation for the
was 108.2 Ϯ 1.6% (n ϭ 18). Acceptable recov-
five assays was 6.7%. The manufacturer’s re-
ery of exogenous analyte (within 90–110%) and
ported cross-reactivity of DSL testosterone an-
demonstration of parallelism suggested no sam-
tisera was 100% with testosterone, 6.6% with
ple matrix effects (Jeffcoate 1981, Grotjan and
5␣-dihydrotestosterone, 2.2% with 5-andro-
Keel 1996, O’Fegan 2000). Extracts from fecal
stane-3, 17-diol, 1.8% with 11-oxotestoste-
For each of the three response variables (es-
trogen, testosterone, E:T), the sampling subglo-bal model adequately fit the data (Table 1). Among the four sampling models, the modelESTR
thus was included with second-stage models(Table 2). The model TEST
tosterone data and thus was included with sec-ond-stage models (Table 3). Similarly, for the E:T data, the model E:T
ported and was included with second-stage mod-els (Table 4).
SEX*RLHS subglobal model adequately fit thedata, whereas the habitat subglobal model didnot
CORT*DATE subglobal models adequately fitthe estrogens and testosterone data, but did notfit the E:T data (Table 1).
For fecal estrogens, the model ESTRCORT*RLHS
was the best supported, with more than doublethe AIC weight of the nearest competing model
25% of the total observed variation. The nearestcompeting model, ESTR
was also well supported and was within two
⌬AIC units of the best model. Both models in-
dicated that fecal estrogen levels were positively
gens and (B) fecal testosterone for California SpottedOwls. Curves of percent binding of I125 tracer (%B
related to fecal glucocorticoid levels in non-
BoϪ1) versus serially diluted (log-transformed doses of
breeding Spotted Owls and unrelated to fecal
1:2 up to 1:128) low pool (n ϭ 2), medium pool (n ϭ
glucocorticoids in breeding owls. The second
2), and high pool (n ϭ2) fecal extracts from male and
model indicated that fecal estrogen levels were
female free-ranging California Spotted Owls were par-
negatively related to sample mass and initial
allel (test of equal slopes, all P Ͼ 0.3) to total estrogen(log-transformed doses of 5–200 pg mLϪ1) or testos-
storage of the samples in liquid nitrogen.
terone standard curves (log-transformed doses of 0.1–
25 ng mLϪ1). Diamonds: total estrogen or testosterone
standard curve points, circles: serially diluted low fecal
ue, and its AIC weight was more than three
extracts, triangles: serially diluted medium pool fecal
extracts, and stars: serially diluted high pool fecal ex-
times higher than the other models (Table 3).
This model explained 29% of the observed var-iation. The nearest competing models wereTEST
rone, and Ͻ1% for other steroids. The sensitivity
which were also supported and within four
⌬AIC units of the best model. These three mod-
els indicated that male California Spotted Owls
had higher fecal testosterone levels than females
and that fecal testosterone levels decreased as
During the 2001 breeding season, we collected
the mass of fecal samples increased. Addition-
and assayed 142 fecal samples from 65 individ-
ally, two of the models suggested that breeding
ual California Spotted Owls (32 females and 33
owls had higher fecal testosterone levels.
males) for fecal estrogen and fecal testosterone
For E:T, the most supported models were E:
metabolites. We collected an average of 2.2 Ϯ
0.1 (range 1–6) fecal samples per individual
which explained 27% of the total observed var-
Ranking of a priori models estimating fecal estrogens in California Spotted Owls in the north-
central Sierra Nevada, California, April–August 2001. Models were ranked based on Akaike’s InformationCriterion adjusted for small sample size (AICc). ⌬AICc is the difference in AICc between a model and the best-approximating model. AICc weights sum to 1 and indicate the relative likelihood of the current model. Seeappendices for explanation of covariate abbreviations and model hypotheses.
a Maximized log-likelihood value. b Number of model parameters. c The lowest AICc score was 1125.4 for the sampling models and 1097.5 for the overall models.
d Covariates from this model were then included in the overall models.
iation. Both E:T models indicated that E:T ratios
life-history stage of individual owls was corre-
were higher in female California Spotted Owls
(Fig. 2). The nearest competing model was E:T
within two ⌬AIC units of the best model (Table
Our findings indicated the ICN I125 double-anti-
4). All three models indicated that the ratio of
body total estrogens RIA and the DSL I125 dou-
E:T was higher in female Spotted Owls com-
ble-antibody testosterone RIA used in this study
pared to males (Fig. 2) and that E:T ratios were
were effective for quantifying immunoreactive
negatively related to sample mass. Additionally,
estrogen and testosterone metabolites, respec-
two of the models suggested the reproductive
tively, in feces from male and female Spotted
Ranking of a priori models estimating fecal testosterone in California Spotted Owls in the north-
central Sierra Nevada, California, April–August 2001. Models were ranked based on Akaike’s InformationCriterion adjusted for small sample size (AICc). ⌬AICc is the difference in AICc between a model and the best-approximating model. AICc weights sum to 1 and indicate the relative likelihood of the current model. Seeappendices for explanation of covariate abbreviations and model hypotheses.
a Maximized log-likelihood value. b Number of model parameters. c The lowest AICc score was 971.3 for the sampling models and 954.6 for the overall models.
d Covariates from this model were then included in the overall models.
Ranking of a priori models estimating fecal estrogen:testosterone ratios (E:T) in California Spotted
Owls in the north-central Sierra Nevada, California, April–August 2001. Models were ranked based on Akaike’sInformation Criterion adjusted for small sample size (AICc). ⌬AICc is the difference in AICc between a modeland the best-approximating model. AICc weights sum to 1 and indicate the relative likelihood of the currentmodel. See appendices for explanation of covariate abbreviations and model hypotheses.
a Maximized log-likelihood value. b Number of model parameters. c The lowest AICc score was 559.6 for the sampling models and 529.7 for the overall models.
d Covariates from this model were then included in the overall models.
Owls. The performance characteristics (e.g., par-
ing fecal samples from Spotted Owls of known
allelism, recovery of exogenous analyte, accu-
sex will be needed to verify the usefulness of
racy, precision) of these assays verified that they
this technique and to determine the potential er-
were accurate, precise, and had an appropriate
range of sensitivity. Monfort et al. (1997) suc-
Sampling factors and characteristics of indi-
cessfully used the same total estrogens and tes-
vidual owls influenced reproductive steroid me-
tosterone assays to quantify fecal estrogen and
tabolite levels in California Spotted Owl feces
testosterone metabolites, respectively, in African
during the breeding season. The amount of feces
wild dogs (Lycaon pictus). Our findings corrob-
available for analysis influenced the estrogen
orate other studies suggesting the reproductive
and testosterone measurements in Spotted Owls.
condition of captive (Bishop and Hall 1991, Ber-
Consequently, the E:T ratios also were influ-
covitz et al. 1982, Lee et al. 1995) and free-
enced by fecal sample mass. Tempel and Gu-
tie´rrez (2004) found a similar relationship be-
Rounce 1995) birds can be monitored using fe-
tween fecal mass and fecal glucocorticoid levels
cal reproductive steroid metabolite assays. Ad-
in California Spotted Owls. Samples of small
ditionally, studies of domestic geese (Anser an-
mass (e.g., 0.02 g dry weight) might have been
ser; Hirschenhauser et al. 2000) and domestic
biased high. Although the reason for this bias
fowl (Gallus domesticus; Cockrem and Rounce
was unknown, we suspected the extraction effi-
1994) have demonstrated that fecal levels of re-
ciency of steroid metabolites from the fecal ma-
productive steroid metabolites were related to
terial may have been higher with very small fe-
cal samples. Researchers should be aware that
Previous studies have demonstrated that fecal
small samples might provide spurious results.
reproductive steroid analyses can be used to de-
Thus, we suggest that in future studies of fecal
termine the sex of individuals from monomor-
reproductive steroids in Spotted Owls, research-
phic bird species (Bercovitz et al. 1978, Stavy
ers decide a priori whether to exclude very
et al. 1979). The E:T ratio has been shown to
small (e.g., Ͻ0.02 g dry weight) fecal samples
be particularly useful. Our findings suggest E:T
from analysis. Additionally, the uric acid portion
ratios from California Spotted Owl feces might
of the samples, which might vary among sam-
be used to determine the sex of the owl. An E:
ples, could have influenced our results.
T ratio of 2.9 or higher indicated the owl was a
Fecal samples with high levels of fecal estro-
female, whereas an E:T ratio of 1.6 or less in-
gen metabolites also had high levels of fecal glu-
dicated the owl was a male. Further studies us-
cocorticoids. This finding was unexpected, as
coids (such as metabolizing energy reserves) un-related to stress responses.
While we observed that variability in sex ste-
roids was related to sampling and sex of thebirds, our models explained relatively little var-iation. Our results suggested a more complex re-lationship among Spotted Owl sex steroids andfeatures of the environment than could be ex-plained by our a priori models. As with otherfecal steroid techniques (e.g., glucocorticoid as-says), it will be important to understand the bi-otic and abiotic factors influencing these hor-mones before the real utility of these techniquescan be realized. Basic studies that examine in-dividual variation and between-year variation inreproductive steroids among birds in differentphysical conditions and life-history stages willbe needed to understand variability in fecal re-productive steroid metabolites. Further work isneeded to determine the influence of fecal sam-ple collection and processing on fecal reproduc-tive steroid measurements. In addition, the in-terrelationships between acute and chronic stressresponses and reproductive hormone levels needto be thoroughly examined. These issues will ne-cessitate the study of birds under controlled con-ditions where environmental conditions (e.g.,food availability) and sample handling are al-tered.
In summary, we validated radioimmunoassay
procedures that quantify estrogen and testoster-one metabolites in Spotted Owl feces. Nonin-vasive monitoring of reproductive steroid me-
Estrogen:testosterone ratios of California
tabolite levels in feces, used in combination with
Spotted Owls in the central Sierra Nevada, California,
demographic information (e.g., reproductive in-
during April–August 2001. Bars represent 95% CI. (A)adult males and females, (B) breeding and nonbreeding
dices), may provide an effective tool for exam-
owls, and (C) male and female breeding (filled circles)
ining the effects of various naturally occurring
and nonbreeding (unfilled circles) owls.
and anthropogenic influences on the reproduc-tive aspects of Spotted Owl populations. Futureresearch examining how fecal reproductive ste-
chronically high corticosterone levels have been
roid levels may vary between male and female
shown to suppress reproduction in birds (Wing-
Spotted Owls during early stages of the breeding
field 1988, Wingfield et al. 1998). Sampling fac-
cycle (e.g., courtship), during the nonbreeding
tors (e.g., sample mass) also influence fecal glu-
season, in juvenile birds, and in other owl pop-
cocorticoid and fecal reproductive steroid con-
ulations inhabiting areas of differing habitat
centrations (Tempel and Gutie´rrez 2004; this
quality are necessary. Additional research is
study). We suspected one or more factors that
needed to assess fundamental issues related to
elevated fecal glucocorticoid levels also elevated
fecal reproductive steroid metabolites, such as
fecal estrogen levels in the same individuals.
sampling issues and understanding normal pro-
That is, birds in breeding status might have a
files in birds of various conditions and life his-
concomitant increase in stress hormone secre-
tory stages, before the technique can be applied
tion related to permissive actions of glucocorti-
for collecting and analyzing demographic data onthe Northern Spotted Owl. Studies in Avian Bi-
Financial and logistical support for this project was pro-
vided by Region 5, U.S. Forest Service (contract
FRANKLIN, A. B., D. R. ANDERSON, R. J. GUTIE´RREZ,
#USDA-FS/53-9158-02-EC06 to RJG), Region 1, U.S.
AND K. P. BURNHAM. 2000. Climate, habitat qual-
Fish and Wildlife Service (service order #101811M663
ity, and fitness in Northern Spotted Owl popula-
to RJG), a University of Missouri (MU) Life Science
tions in northwestern California. Ecological
Mission Enhancement Postdoctoral Fellowship, a MU
Research Board Grant, the MU Department of Fish-
FRANKLIN, A. B., R. J. GUTIE´RREZ, J. D. NICHOLS, M.
eries and Wildlife Sciences, and the University of Min-
E. SEAMANS, G. C. WHITE, G. S. ZIMMERMAN, J.
nesota. Fecal hormone metabolite assays were con-
E. HINES, T. E. MUNTON, W. S. LAHAYE, J. A.
ducted in the Wildlife Physiology Laboratory in the
BLAKESLEY, G. N. STEGER, B. R. NOON, D. W. H.
MU Department of Fisheries and Wildlife Sciences.
SHAW, J. J. KEANE, T. L. MCDONALD, AND S. BRIT-
We thank M. Larson for assistance with the statistical
TING. 2004. Population dynamics of the California
analyses. G. Zimmerman provided insightful com-
Spotted Owl (Strix occidentalis occidentalis): a
ments that greatly improved the manuscript. This re-
meta-analysis. Ornithological Monograph 54.
search was approved by the University of Minnesota
GROTJAN, H. E., AND B. A. KEEL. 1996. Data interpre-
Animal Care and Use Committee (Animal Subjects
tation and quality control, p. 51–93. In E. P. Dia-
mandis and T. K. Christopoulos [EDS.], Immuno-assay. Academic Press, New York.
GUTIE´RREZ, R. J., J. VERNER, K. S. MCKELVEY, B. R.
NOON, G. N. STEGER, D. R. CALL, W. S. LAHAYE,
BARROWCLOUGH, G. F., R. J. GUTIE´RREZ, AND J. G.
B. B. BINGHAM, AND J. S. SENSE. 1992. Habitat
GROTH. 1999. Phylogeography of Spotted Owl
relations of the California Spotted Owl, p. 79–98.
(Strix occidentalis) populations based on mito-
In J. Verner, K. S. McKelvey, B. R. Noon, R. J.
chondrial DNA sequences: gene flow, genetic
Gutie´rrez, G. I. Gould, Jr., and T. W. Beck [EDS.],
structure, and a novel biogeographic pattern. Evo-
The California Spotted Owl: a technical assess-
ment of its current status. USDA Forest Service
BERCOVITZ, A. B., J. COLLINS, P. PRICE, AND D. TUTTLE.
General Technical Report PSW-GTR-133.
1982. Noninvasive assessment of seasonal hor-
GUTIE´RREZ, R. J., A. B. FRANKLIN, AND W. S. LAHAYE.
mone profile in captive Bald Eagles (Haliaeetus
1995. Spotted Owl (Strix occidentalis), p. 1–28. leucocephalus). Zoo Biology 1:111–117. In A. Poole and F. Gill [EDS.], The birds of North
BERCOVITZ, A. B., N. M. CZEKALA, AND B. L. LASLEY.
America, No. 179. The Academy of Natural Sci-
1978. A new method of sex determination in
ences, Philadelphia, PA, and The American Or-
monomorphic birds. Journal of Zoo and Animal
nithologists’ Union, Washington, DC.
HIRSCHENHAUSER, K., E. MO¨STL, P. PE´CZELY, B. WALL-
BISHOP, C. M., AND M. R. HALL. 1991. Non-invasive
NER, J. DITTAMI, AND K. KOTRSCHAL. 2000. Sea-
monitoring of avian reproduction by simplified
sonal relationships between plasma and fecal tes-
faecal steroid analysis. Journal of Zoology (Lon-
tosterone in response to GnRH in domestic gan-
ders. General and Comparative Endocrinology
BURNHAM, K. P., D. R. ANDERSON, AND G. C. WHITE.
1996. Meta-analysis of vital rates of the Northern
JEFFCOATE, S. L. 1981. Efficiency and effectiveness in
Spotted Owl. Studies in Avian Biology 17:92–
the endocrine laboratory. Academic Press, San Di-
BURNHAM, K. P., AND D. R. ANDERSON. 1998. Model
KOFUJI, H., M. KANDA, AND T. OISHI. 1993. Breeding
selection and inference: a practical information-
cycles and fecal gonadal steroids in the Brown
theoretic approach. Springer-Verlag, New York.
Dipper (Cinclus pallasii). General and Compara-
COCKREM, J. F., AND J. R. ROUNCE. 1994. Faecal mea-
surements of oestradiol and testosterone allow the
KU¨CHLER, A. W. 1977. The map of natural vegetation
non-invasive estimation of plasma steroid concen-
of California, p. 908–938. In M. G. Barbour and
trations in the domestic fowl. British Poultry Sci-
J. Major [EDS.], Terrestrial vegetation of Califor-
COCKREM, J. F., AND J. R. ROUNCE. 1995. Non-invasive
LEE, J. V., C. S. WHALING, B. L. LASLEY, AND P. MAR-
assessment of the annual gonadal cycle in free-
LER. 1995. Validation of an enzyme immunoassay
living Kakapo (Strigops habroptilus) using fecal
for measurement of excreted estrogen and testos-
steroid measurements. Auk 112:253–257.
terone metabolites in the White-crowned Sparrow
FORSMAN, E. D. 1983. Methods and materials for lo-
(Zonotrichia leucophrys oriantha). Zoo Biology
cating and studying Spotted Owls. USDA Forest
Service General Technical Report PNW-GTR-
LE MAHO, Y., H. KARMANN, D. BRIOT, AND Y. HAN-
DRICH. 1992. Stress in birds due to routine han-
FORSMAN, E. D., E. C. MESLOW, AND H. M. WIGHT.
dling and a technique to avoid it. American Jour-
1984. Distribution and biology of the Spotted Owl
LITTELL, R. C., G. A. MILLIKEN, W. W. STROUP, AND
FRANKLIN, A. B., D. R. ANDERSON, E. D. FORSMAN, K.
R. D. WOLFINGER. 1996. SAS system for mixed
P. BURNHAM, AND F. F. WAGNER. 1996. Methods
models. SAS Institute Inc., Cary, NC.
MONFORT, S. L., S. K. WASSER, K. L. MASHBURN, M.
TEMPEL, D. A., AND R. J. GUTIE´RREZ. 2004. Factors
BURKE, B. A. BREWER, AND S. R. CREEL. 1997.
related to fecal corticosterone levels in California
Steroid metabolism and validation of noninvasive
Spotted Owls: implications for assessing chronic
monitoring in the African wild dog (Lycaon pic-
stress. Conservation Biology 18:538–547. tus). Zoo Biology 16:533–548.
U.S. DEPARTMENT OF THE INTERIOR. 1990. Endangered
NETER, J., W. WASSERMAN, AND M. H. KUTNER. 1990.
and threatened wildlife and plants; determination
Applied linear statistical models. 3rd ed. Irwin
of threatened status for the Northern Spotted Owl.
NOON, B. R., AND A. B. FRANKLIN. 2002. Scientific
U.S. DEPARTMENT OF THE INTERIOR. 1993. Endangered
research and the Spotted Owl (Strix occidentalis):
and threatened wildlife and plants: final rule to list
opportunities for major contributions to avian
the Mexican Spotted Owl as a threatened species.
population ecology. Auk 119:311–320.
O’FEGAN, P. O. 2000. Validation, p. 211–238. In J. P.
EPARTMENT OF THE INTERIOR. 2003. 12-month
finding for a petition to list the California Spotted
Owl (Strix occidentalis occidentalis). Federal
EERY, M. Z., R. J. GUTIE´RREZ, AND M. E. SEAMANS.
1999. Habitat composition and configuration
around Mexican Spotted Owl nest and roost sites
VERNER, J., K. S. MCKELVEY, B. R. NOON, R. J. GU-
in the Tularosa Mountains, New Mexico. Journal
´ RREZ, G. I. GOULD JR., AND T. W. BECK. 1992.
Assessment of the current status of the California
RUNDEL, P. W., D. J. PARSONS, AND D. T. GORDON.
Spotted Owl, with recommendations for manage-
1977. Montane and subalpine vegetation of the
ment, p. 3–26. In J. Verner, K. S. McKelvey, B.
Sierra Nevada and Cascade ranges, p. 559–599.
R. Noon, R. J. Gutie´rrez, G. I. Gould, Jr., and T. In M. G. Barbour and J. Major [EDS.], Terrestrial
W. Beck [EDS.],The California Spotted Owl: a
vegetation of California. J. Wiley and Sons, New
technical assessment of its current status. USDA
Forest Service General Technical Report PSW-
SAS INSTITUTE. 1985. SAS user’s guide: statistics. Ver-
sion 5. SAS Institute Inc., Cary, NC.
WASSER, S. K., K. BEVIS, G. KING, AND E. HANSON.
SCHWARZENBERGER, F., E. MO¨STL, E. BAMBERG, J. PAM-
1997. Noninvasive physiological measures of dis-
MER, AND O. SCHMEHLIK. 1991. Concentrations of
turbance in the Northern Spotted Owl. Conser-
progestagens and oestrogens in the faeces of preg-
nant Lipizzan, trotter and thoroughbred mares.
WINGFIELD, J. C. 1988. Changes in reproductive func-
Journal of Reproduction and Fertility Supplement
tion of free-living birds in direct response to en-
vironmental perturbations, p. 121–148. In J. H.
EAMANS, M. E., R. J. GUTIE´RREZ, C. A. MOEN, AND
Stetson [ED.], Processing of environmental infor-
the central Sierra Nevada. Journal of Wildlife
mation in vertebrates. Springer-Verlag, Berlin.
WINGFIELD, J. C., C. BREUNER, J. JACOBS, S. LYNN, M.
STAVY, M., D. GILBERT, AND R. D. MARTIN. 1979. Rou-
RAMENOFSKY, AND R. RICHARDSON. 1998. Ecolog-
tine determination of sex in monomorphic species
ical basis of hormone-behavior interactions: the
using fecal steroid analysis. International Zoo
emergency life history stage. American Zoologist
Covariates used in a priori models to estimate fecal estrogens, fecal testosterone, and fecal
estrogens:testosterone ratios in California Spotted Owls in the north-central Sierra Nevada, California, April–August 2001.
Storage method (same initially stored in liquid N2 or on ice)
Mass of dried sample analyzed (to the nearest 0.001 g)
Reproductive life-history stage (breeding or nonbreeding)
Fecal glucocortiocoid metabolite level (ng gϪ1) estimated from the same sample
Amount of interior Spotted Owl habitat (Ͼ100 m from an edge) within the
Number of discrete patches of Spotted Owl habitat within the owl’s territory
Description, representation, and predicted directions of a priori models to estimate fecal estro-
gens, fecal testosterone, and fecal estrogens:testosterone ratios in California Spotted Owls in the north-centralSierra Nevada, California, April–August 2001. Each model was evaluated for each of the three response variables(estrogens, testosterone, and E:T).
reproductive life-histo-ry stage of the bird
are related to repro-ductive life-historystage of the bird
the number of patcheswithin an owl’s terri-tory
the amount of corearea within an owl’sterritory
and the amount ofcore area within anowl’s territory
are related to theamount of core areawithin its territory
a Expected direction of coefficients, given that the hypothesized model is the most supported.
SQUASH PROFILES EET A SQUASH PLAYER Kimberley Bessell Age: 15 Date of Birth: 1st March 1989 School: Werribee Grange Secondary College - Year 10 Place of Residence: Hoppers Crossing Favourite Foods: Lasagne, Ravioli Hobbies: Music, movies and hanging around Idol: Sarah Fitz-Gerald - 5 times World Open Squash Champion Greatest Influences: Parents and her
1.- INSCRIPCION 2.- ORGANIZACIÓN DE CARRERAS. (PERSONAL) 3.- FRECUENCIAS Y CAMBIOS DE FRECUENCIAS 4.- EMISORAS Y CONTROL DE EMISORAS 5.- FORMATO DE CARRERA. Entrenamientos Inspección Técnica Reunión de pilotos Grupos clasificatorias Sub finales y Final Procedimiento cuentavueltas 6.- SANCIONES 7.- RECLAMACIONES 8.- BANDERAS 1.- INSCRIPCIÓN 1. El piloto deberá formalizar su inscripción y
 For each of the three response variables (es-
trogen, testosterone, E:T), the sampling subglo-bal model adequately fit the data (Table 1).
For each of the three response variables (es-
trogen, testosterone, E:T), the sampling subglo-bal model adequately fit the data (Table 1).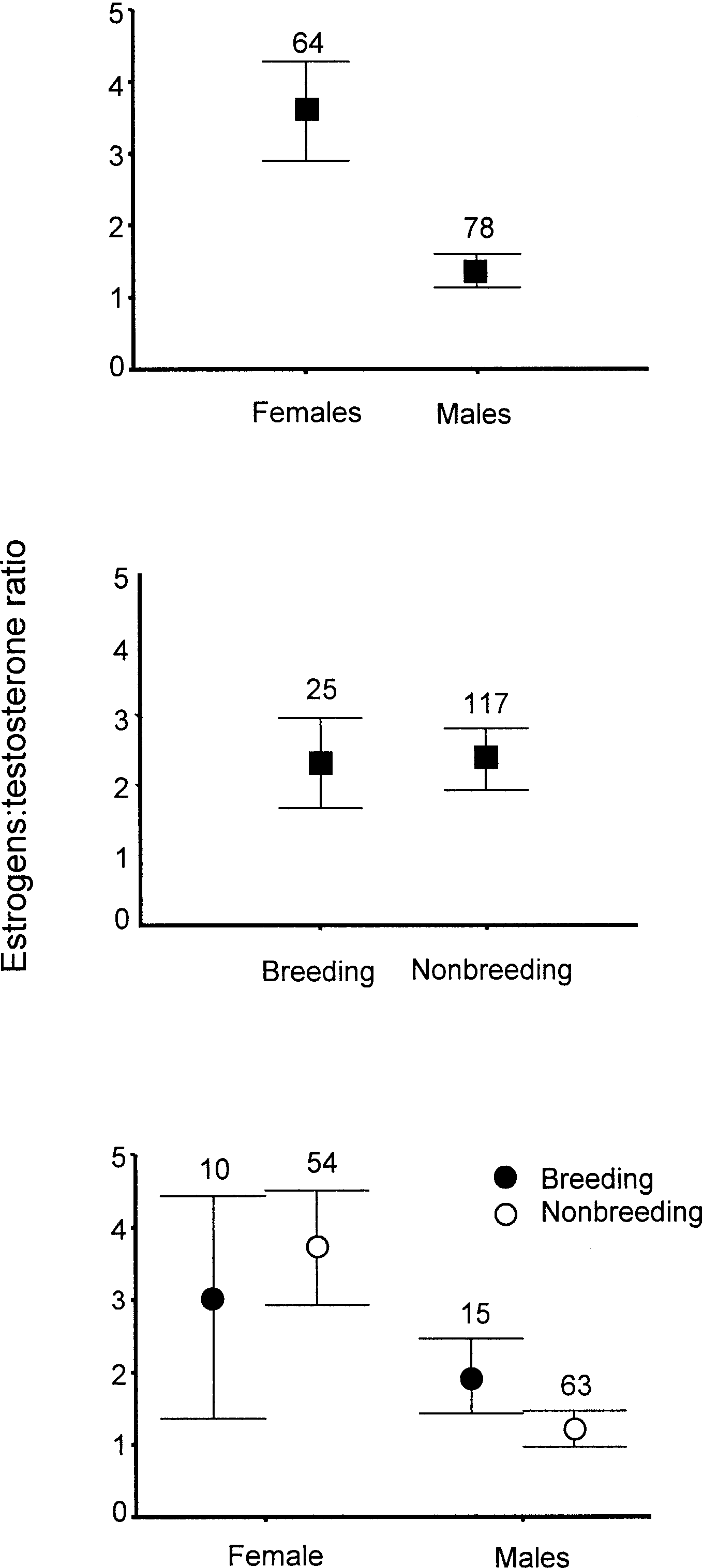 coids (such as metabolizing energy reserves) un-related to stress responses.
coids (such as metabolizing energy reserves) un-related to stress responses.